Principalii factori care influențează productivitatea fitoplanctonului. Biomasă, producție primară de fitoplancton și clorofilă. Ciclul de producere a fitoplanctonului
Ciclul de producere a fitoplanctonului
Principalii factori care determină producția primară de fitoplancton sunt grosimea stratului fotic, aprovizionarea cu fitoplancton cu nutrienți și durata ciclului de producție.
În general, acești parametri depind de condițiile climatice și hidrologice specifice fiecărei regiuni și se manifestă în momentul începerii dezvoltării primăverii, care este strâns legat de regimul de urme și de sfârșitul ciclului de producție; în plus, acești parametri sunt determinați de aportul de nutrienți formați în stratul fotic la începutul dezvoltării primăverii și determinați de scurgerea râului și de adâncimea amestecării convective de iarnă, difuzia turbulentă verticală de vară, cantitatea de energie solară (în funcție de rândul ei). privind perioada anului și factorii climatici), precum și caracteristicile structurale și funcționale ale fitoplanctonului.
Fitoplanctonul Mării Albe este planctonul sărăcit calitativ al Mării Barents, cu un amestec mare de forme epifitice „litorale” și de apă dulce (Kokin, Koltsova, 1972). În Marea Albă, comparativ cu partea de sud a Mării Barents, numărul speciilor este dominat de diatomee iubitoare de frig, iar diversitatea speciilor de peridini este redusă. Tivul conținutului de alge din compoziția fitoplanctonului (63%) a Mării Albe este aproape de mările epicontinentale ale arctice înalte precum Kara (67%) și Laptev (64%), ceea ce indică încă o dată natura sa arctică. Formarea compoziției calitative și cantitative a fitoplanctonului în diferite anotimpuri biologice are loc sub influența factorilor de mediu atât biotici, cât și abiotici.
Schimbările în indicatorii structurali și funcționali ai comunității în timpul ciclului de producție pot fi urmărite prin exemplul analizei anotimpurilor biologice din Golful Kandalaksha în 1970 (Fedorov și Bobrov, 1977, Bobrov, 1982).
În total, în perioada de observație de la 15 aprilie până la 26 octombrie 1970, au fost instalate 51 de stații (de două ori pe săptămână). Produsele au fost determinate folosind metoda radiocarbonului baloanelor la orizonturi de 0,2,5, 5, 10, 25, 50, 75 m și la fund. Radiația solară totală a fost măsurată cu un piranometru Janyshevsky într-o modificare subacvatică. Concomitent cu experimentele de determinare a producției primare din orizonturile corespunzătoare, s-au efectuat colecții batometrice cantitative de fitoplancton și studiul unor parametri hidrologici și hidrochimici. La calcularea eficienței procesului fotosintetic, aceste măsurători ale producției primare au fost exprimate în unități energetice, pe baza următorului calcul: se consumă 112 calorii de energie luminoasă pentru a fixa 1 mol de CO2, iar această valoare corespunde la 9,36 cal/mgC.
Trecerea de la numărul de indivizi fitoplancton la caracteristicile de greutate s-a făcut după formula:
Formula a fost obținută ținând cont de faptul că biomasa uscată de alge conține 35% cenușă (Fedorov și colab., 1974).
Valorile cantităților zilnice de radiații în timpul sezonului de vegetație din 1970 au variat între 20,6 și 2300 cal/(cm") zi. Creșterea radiației solare care a pătruns în BOAy în primele zece zile ale lunii mai până la 100,6-181,4 cal/( cm" zi) coincide cu ruperea stratului de gheață. O ușoară scădere a valorilor energiei solare totale a fost cauzată de condițiile meteorologice.Îmbunătățirea condițiilor meteorologice și creșterea duratei orelor de lumină au dus la coincidența practică a maximului luminos (2300 cal/(cm. zi)) cu ziua calendaristică a solstițiului de vară.Scăderea ulterioară a cantităților zilnice de radiație solară are un caracter destul de neted și este cauzată de o scădere a duratei LEHI HEM CBPTOBOTO IH H H BECOTE CONHL kl Până la sfârșitul sezonului de vegetație, cantitatea de energie solară scade la 20 cal/(cm. zi).
Astfel, pentru întreaga perioadă de observare, care acoperă 183 de zile, valoarea energetică a fost exprimată ca valoare totală de 22,8 kcal/cm Energia necontabilizată a radiației solare penetrante în perioada de toamnă a „izvorului subglaciar” a fost obținută prin extrapolare și a fost însumată până la 22 kcal/cm.” În total, 25 kcal/cm au pătruns în rezervor în timpul sezonului." Această din urmă valoare poate fi considerată o sumă anuală, deoarece în lunile de iarnă cantitatea de energie de radiație solară care pătrunde sub stratul de gheață este nesemnificativă. Studiul distribuției. a energiei solare în coloana de apă în perioada de vegetație a făcut posibilă calcularea coeficientului de absorbție a luminii cu apă k=0,37-002.
Temperatura din orizontul de suprafață în timpul sezonului de vegetație a variat între -1,54 și 15,65 °C. Fluctuațiile de temperatură s-au atenuat cu adâncimea. Perioada de primăvară după spargerea gheții este caracterizată de o încălcare a homotermiei și de stabilirea unei stratificări clar definite. Zona de salt de temperatură, cu o creștere a energiei solare penetrante și încălzirea corespunzătoare a VM, s-a mutat treptat de la suprafață la straturile mai profunde. În perioada de maximă energie solară, termoclinul a fost situat la o adâncime de 25 m și a rămas la acest orizont până la sfârșitul lunii septembrie. În octombrie, saltul de temperatură a fost localizat mai adânc și s-a dezvăluit o tendință spre homeotermie.
În tabel Tabelul 5.1 prezintă câteva caracteristici abiotice ale Mării Albe, obținute pe baza analizei LHHaMHK.H sezoniere ale acestora.
Analiza dezvoltării vegetative a planctonului și a producției sale HEIM Xapa KTepH cTH K face posibilă identificarea și stabilirea duratei principalelor perioade de vegetație, numite biologice de primăvară, vară și toamnă de V.G.Bogorov (1938). Marea Albă este caracterizată de toate cele trei perioade. Durata lor pentru Golful Kandalaksha în 1970 a fost de 48, 92 și, respectiv, 43 de zile. 
Rezultatele care caracterizează valorile absolute ale procesului de producție în diferite perioade ale sezonului de vegetație sunt prezentate în tabel. 5.2. Coeficientul P/Sent a fost calculat ca raportul dintre producția primară și biomasă (exprimată în carbon) pentru Bice TO CTO,JI (5a B OIE bil. 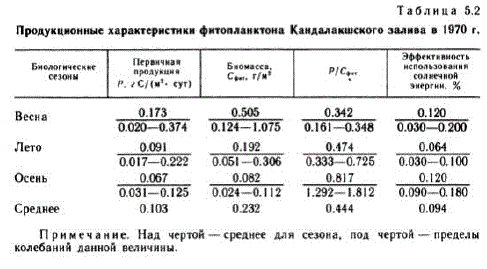
Notă. Deasupra liniei este media pentru sezon, sub linie este intervalul de fluctuații ale unei anumite valori.
Începutul primăverii biologice s-a remarcat în primele zece zile ale lunii aprilie Punctul culminant subglaciar de primăvară (Fedorov, 1970a; Kokin, Koltsova, 1972) durează aproximativ 2 săptămâni și se caracterizează printr-un coeficient P/Cphi stabil. Distribuția econichelor de alge corespunde celui de-al doilea model MacArthur (Kokin și Koltsova, 1972).
Izbucnirea de primăvară a abundenței de biomasă și alge începe sub gheață și atinge un maxim după ce gheața se rupe. Fitoplanctonul este reprezentat de BH La MH JULHETOMOBOTO KOM TIJEKCE: Fragilaria oceanica, Chaetoceros holsa ficus, Nitzschia frigida, Maticula septentrionalia. Cea mai abundentă specie a fost F. oceapisa. Stratul fotic este îmbogățit cu substanțe nutritive și conține 86% din biomasa totală de fitoplancton, care, ca și rata fotosintezei, în această perioadă atinge valoarea maximă pentru întregul sezon de vegetație (vezi Tabelul 5.2). Consumul de alge de către zooplancton (Konoplya, 1973) duce la o scădere a biomasei de alge și la o scădere a competiției alimentare pentru nutrienți, în timp ce distribuția algelor corespunde deja primului model MacArthur.
Maximul de vară al dezvoltării fitoplanctonului (primul), cauzat de creșterea numărului de diatomee, după o scădere a numărului acestora, este „preluat” de algele peridiniu, formând al doilea maxim de vară. De la mijlocul lunii iulie până la sfârșitul lunii august, punctul culminant al verii este stabilit cu un raport stabil P/C-fit. Formele dominante ale verii biologice sunt Sceletopema costalum și speciile din genul Chaetoceros (în principal Ch. compostis). Productia primara in aceasta perioada se caracterizeaza printr-o valoare medie de 0,091 g C/(m", zi), iar biomasa - 0,192 g C/m". Se dezvoltă competiția trofică pentru nutrienți,
Focarul de toamnă al fitoplanctonului este slab exprimat. Se observă de la sfârșitul lunii august până la mijlocul lunii septembrie și creșterea biomasei ee, Rb Ae/y!"b, "pyAhQ. Doar o creștere a producției primare la 0,125 g C/(m", zi) și a coeficientului P/Seite (până la 1812) pe 20 septembrie indică o activare pe termen scurt a fitoplanctonului. În această perioadă, dezvoltarea Dinophyes powegica. , o dominantă iubitoare de frig a toamnei biologice, se observă ( Fedorov, 1969).Perioada de succesiune a toamnei duce ecosistemul la un apogeu pe termen scurt, care durează până la sfârșitul lunii septembrie.O scădere suplimentară a temperaturii și radiației solare, precum si o crestere a stratului de amestec datorita perturbarii saltului de densitate duce la o redistribuire a biomasei fitoplanctonului in profunzime si la predominarea proceselor de distrugere asupra produselor.
În general, un astfel de curs al ciclului de producție a fitoplanctonului poate fi de așteptat în alte zone ale mării. Posibilele diferențe vor fi exprimate în momentul începerii și sfârșitului sezonului de vegetație a fitoplanctonului, precum și în unele diferențe în compoziția calitativă a speciilor de alge dominante (Zhitina, 1981), care vor afecta fără îndoială rezultatele proceselor de producție a fitoplanctonului. în aceste părţi ale zonei de apă.
Datele din studiile ciclului de producție a fitoplanctonului din 1970 au făcut posibilă clarificarea clasificării troficității Mării Albe. Ca o caracteristică suplimentară, a fost utilizată eficiența utilizării energiei solare de către fitoplancton (vezi Tabelul 5.2), care este o caracteristică integrală a stării fitoplanctonului, în funcție de un set de condiții de mediu și neprezentând o legătură directă cu biomasa fitoplanctonului. (Fedorov, Bobrov, 1977).
Dacă comparăm valorile calculate ale principalelor caracteristici de producție ale fitoplanctonului cu datele cunoscute pentru regiunile oceanice cu productivitate diferită (Sorokin, 1973), putem concluziona că Golful Kandalaksha aparține unor zone neproductive cu un nivel de producție de fitoplancton de 0,05-0,10 g. C/(m* - zi) și o eficiență a utilizării energiei solare de 0,04%, caracteristică apelor oligotrofe. În consecință, Golful Kandalaksha, conform clasificării O.I. Koblenz-Mishke (Koblenz-Mishke și colab., 1970), poate fi clasificat ca clasa trofică 1 cu foarte puțin amestec de ape adânci.
Tabelele 6.7-6.9 sintetizează rezultatele măsurătorilor producției primare în apele diferitelor recife. După cum se poate observa din tabele, producția primară variază foarte mult chiar și în apele aceluiași recif. Cu toate acestea, aceste din urmă fluctuații sunt destul de naturale și sunt asociate în primul rând cu modificări ale concentrației de fitoplancton în secțiuni din zonele de mică adâncime ale recifului spre centrul lagunei.
În lagunele atolilor cu producție primară scăzută (atolurile Insulei Lau și Insulele Marshall, Insulele Tuamotu), limita inferioară (2-4 mg C/m 3 pe zi) se apropie de valorile sale în oligotrofic. ape tropicale ale curenților alizei care le spală, unde de obicei este de 1-5 mg C/m 3 pe zi (Sorokin, 1971a). Cu toate acestea, chiar și în astfel de atoli, în apele lagunelor acestora sunt posibile izbucniri sezoniere de dezvoltare a fitoplanctonului, care pot atinge intensitatea unei „înfloriri” (Sournia, Ricard, 1976a). După cum se poate observa din datele din tabel. 6.7, în lagunele de atoli, chiar și cele spălate de ape tropicale oligotrofe cu niveluri extrem de scăzute de dezvoltare a fitoplanctonului, producția primară corespunde cel mai adesea nivelului mediu și uneori superior al productivității fitoplanctonului în bazinele marine mezotrofice (15-50 mg C/m 3 per zi). În laguna marelui recif inel Cargados-Carajos, situată în miezul apelor foarte neproductive ale curentului sudic al alizei din Oceanul Indian, producția primară a atins 250 mg C/m 3 pe zi cu o biomasă de alge (în principal diatomee). ) mai mare de 5,2 g/m 3 (Tabelul 6.10). Producția ridicată (70-100 mg C/m3 pe zi) a fost înregistrată într-o lagună închisă a atolului Fanning (Insulele Lau) în timpul perioadei de dezvoltare a dinoflagelatelor și algelor cocoide albastru-verzi (Gordon și colab., 1971). Acest atol este situat și în apele oligotrofe ale curentului alizez. Astfel, chiar și în lagunele recifelor inelare individuale situate în curgerea apelor oligotrofe pot exista comunități de fitoplancton destul de productive, a căror producție și biomasă sunt cu 1-2 ordine de mărime mai mari decât în apele oceanice care le spală.
Judecând după rezultatele puținelor măsurători disponibile, concentrațiile de clorofilă a fotosintetic activă în apele lagunei ale recifelor inelare ale oceanului deschis sunt în medie de 5-10 ori mai mari decât în apele oceanice din jur. Conform măsurătorilor unice, acesta a variat în lagunele diferitelor recife în intervalul de la 0,2 la 1 mg/m3. În laguna atolului Takapoto, conform rezultatelor observațiilor pe termen lung (Sougnia, Ricard, 1976a), concentrația de clorofilă a variat de la 0,26 la 0,77 mg/m 3 și a variat considerabil în diferite părți ale lagunei în funcție de intensitatea pătrunderii în apele oceanului în timpul mareelor înalte.
O contribuție semnificativă la studiul fitoplanctonului și al produselor sale au avut-o observațiile sezoniere efectuate în apele recifelor de barieră și de coastă din apropierea insulelor mari, unde au fost înființate stații biologice pentru a studia viața recifelor și a zonei neritice adiacente a oceanului. Dintre astfel de recife bine studiate, trebuie menționate recifele din largul insulei Nosi Be și din largul Tulear (Madagascar), reciful hawaian din largul insulei Oahu, Marea Barieră de Corali din Australia, recifele din Waprao, Tiaura și Faaa pe insulele Tahiti și Moorea, recifele din centrul Vietnamului lângă Nha Trang. Rezultatele măsurătorilor producției primare și a biomasei fitoplanctonului a recifelor de barieră și de coastă sunt rezumate în tabel. 6.8, 6.9, 6.10: Datele prezentate în ele variază foarte mult pentru aproape toate recifele studiate. Ele reflectă măsurători regulate ale producției primare în diferite zone ale aceluiași recif. În același timp, cele mai mici valori ale sale (precum și cele mai mici valori ale biomasei fitoplanctonului: Fig. 6.10) corespund zonelor de apă puțin adâncă ale recifului, unde apa este în contact intens cu populația de fund și flora algelor. a recifului. În aceste zone (plate, recife petice, litoral) fitoplanctonul, de regulă, este epuizat și producția sa scade brusc aici. Valorile maxime ale producției de fitoplancton și ale biomasei prezentate în aceste tabele pe fiecare recif corespund zonelor lagunei sau pantei exterioare a recifului marginal sau de coastă cu adâncimi mai mari de 15-20 m, unde apa nu este în contact direct cu populația de fund a recifului. Valorile producției primare în aceste zone, de regulă, sunt semnificativ mai mari decât în apele mării deschise adiacente. În medie, ele sunt mai mari decât în apa lagunelor atolilor și recifelor inelare ale oceanului deschis.
Valori ridicate ale producției primare, atingând nivelul apelor eutrofice, s-au remarcat în apele unui număr de recife de coastă și de barieră ale Oceanului Indian, cum ar fi recifele din Madagascar, Insulele Seychelles și Insula Socotra, precum și în laguna recifului de barieră B. în perioadele de „înflorire” al algelor albastre-verzi (vezi Fig. 6.9). Producția zilnică aici a ajuns la 50-300 mg C/m 3 pe zi în stratul superior de apă, sau 0,7-2,8 g C/m 3 . Mai mult, în apele recifelor de barieră ale insulelor spălate de apele oceanice oligotrofe (Tangi, Oahu, Lizard), producția primară în lagune poate ajunge la 50-60 mg C/m 2, sau 0,4-0,7 g de carbon pe 1 m 2 zi. , care corespunde nivelului superior de productivitate al apelor mezotrofe. În zonele de recif ale coastei Vietnamului Central și insulele sale din Marea Chinei de Sud (Insulele Thu) și în Golful Thailandei (Insulele Nam Zu) (a se vedea Tabelul 6.8, Fig. 6.10), valorile fitoplanctonului primar producția a fost ridicată, în ciuda faptului că s-au făcut observații în perioada de minim sezonier de fitoplancton (Sorokin și colab., 1982). Cele mai mari valori ale producției primare, de regulă, au fost observate în apele de la marginea exterioară a recifului, la o distanță de 10-100 m de acesta, peste adâncimi de 5-15 m (vezi Fig. 6.10). De obicei depăşea 30-40 mg C/m 3 pe zi. În secțiunile individuale din această zonă, în 1984, s-a înregistrat producție primară peste 100 mg C/m 3 pe zi.
Informațiile despre biomasa fitoplanctonului de pe recifele de coastă și de barieră sunt încă foarte fragmentare. Cele mai detaliate măsurători ale biomasei fitoplanctonului în timpul sezonului au fost făcute de Angot (1968) pe Insula Nosy Be (Madagascar). Rezultatele acestor măsurători sunt rezumate în Fig. 6.9. În tabel Tabelul 6.10 prezintă datele disponibile în literatura de specialitate cu privire la biomasa fitoplanctonului pe un număr de recife mari din Oceanele Indian și Pacific. După cum se poate observa din tabel, biomasa fitoplanctonului din apele recifurilor de barieră și de coastă poate ajunge la 3-8 g/m3. Cele mai scăzute valori ale sale au fost observate în apele recifelor de coastă și de barieră din partea centrală a Oceanului Pacific, care se aflau într-o stare depresivă (recife de pe insulele Panarora și Paama, Noile Hebride, recif de lângă Insulele Tuvuta, Insulele Lau). ).
Informațiile privind conținutul de clorofilă a din apele recifelor de barieră și de coastă sunt la fel de limitate (Tabelul 6.11). Cele mai detaliate măsurători ale sale au fost efectuate în apele lagunelor insulelor Moorea și Tahiti de către cercetători francezi (Ricard, 1976; Sournia, Ricard, 1976). Aceste măsurători au arătat că concentrația de clorofilă în ele în diferite anotimpuri ale anului variază în intervalul 0,03-0,17 mg/m 3. În laguna recifului hawaiian din largul insulei Oahu, precum și în laguna recifului de barieră B. iarna, concentrația de clorofilă a fost apropiată de cea din apele marine mezotrofice: 0,1-0,3 mg/m3. În perioada de dezvoltare maximă toamnă-vară a fitoplanctonului în laguna recifului de barieră B., precum și în apele recifului din largul insulei Nosy Be (Madagascar), concentrația de clorofilă în apă a fost mai mare cu 1 mg/m 3, care este aproape de conținutul său în apele eutrofice (Angot , 1968). S-a stabilit influenţa resuspendarii întregului strat de sedimente de fund, îmbogăţit în microbentos, în zona digului asupra concentraţiei de clorofilă din apele recifului (Walker, 1981).
După cum sa menționat mai sus, valorile producției primare și ale biomasei fitoplanctonului variază foarte mult în diferite zone ale aceluiași recif. În cele mai multe cazuri, cele mai scăzute valori ale producției de biomasă și fitoplancton sunt caracteristice zonelor de mică adâncime ale recifului și în special platului recifului, unde apa care trece peste recif în timpul mareelor înalte și joase este în cel mai mare contact cu populația de fund a reciful. Motivul scăderii fitoplanctonului în zonele puțin adânci ale platului ar trebui să fie văzut în consumul acestuia de către filtrele de fund ale recifului (Glynn, 1973a). În plus, celulele dnatoms, cum ar fi Nitzschia delicatissima sau celulele ramificate de Chaetoceros, pot fi rănite în apele turbulente ale recifului, în special atunci când se ciocnesc cu suprafața tare a recifului și cu particulele suspendate. În apele din părțile mai adânci ale lagunei sau dincolo de marginea exterioară a recifului, unde contactul apei cu reciful scade, dezvoltarea fitoplanctonului se intensifică datorită eliberării unor substanțe nutritive în comunitățile de fund ale recifului.
În ceea ce privește indicatorii cantitativi ai biomasei și producției sale, precum și în ceea ce privește concentrația de clorofilă, fitoplanctonul din zona recifului este mai bogat decât în apele înconjurătoare ale oceanului deschis (Michel, 1969; Sorokin, 1971; Ricard, 1976; Sournia, Ricard). , 1976a). Figura 6.3 a prezentat o imagine tipică a schimbărilor în producția primară de fitoplancton de-a lungul secțiunilor de la Heron Reef (B. Barrier Reef) până la oceanul deschis. Deja la o distanță de 6-9 km scade cu mai mult de un ordin de mărime.
Transparența apei de-a lungul discului alb în zonele adânci ale recifelor este de obicei destul de mare și are o medie de 10-15 m, deși în unele cazuri poate fi mai mică (5 m pe atolul Canton) și mai mult de 30 m (atolul Majuro). ). Astfel, fotosinteza fitoplanctonului nu este practic limitată de lumină pe toată grosimea lor. În fig. Figura 6.11 prezintă curbele tipice de atenuare pentru radiația incidentă în apele unor recife din Oceanul Indian cu transparență medie (10-18). După cum se poate observa din această figură, fitoplanctonul poate experimenta o oarecare lipsă de lumină doar în partea de jos a lagunei.
Exemple de variabilitate sezonieră a fitoplanctonului sunt prezentate în Fig. 6.9,6.12,6.13. Acestea arată că în diferite anotimpuri ale anului nu numai compoziția sa se modifică (vezi 6.2.1), dar există modificări ale biomasei și activității sale funcționale. Fluctuațiile valorilor producției primare de fitoplancton și ale concentrațiilor de clorofilă de pe recife în timpul sezonului au atins un ordin de mărime (Angot, 1968; Sournia, 1972). Mai mult, chiar și în laguna atolului Takapoto, situată într-o zonă de ape oligotrofe, concentrația de clorofile în timpul maximului sezonier a atins un nivel caracteristic apelor mezotrofe (Sournia și Ricard, 1976). În apele de barieră și recifele de coastă, concentrația sa a fost apropiată de 1 mg/m3, ceea ce corespunde conținutului său în apele eutrofice, iar biomasa de fitoplancton din zona recifului din largul insulei Nosi Be a depășit 3 g/m3. S-au observat fluctuații sezoniere semnificative ale concentrației de clorofilă în apă (de la 0,13 la 0,5-2,5 mg/m 3) în laguna B. a recifului de barieră australian de lângă Townsville (Sammarco, Crenshaw, 1984). Maximul a fost observat în lunile de vară-toamnă din decembrie până în aprilie. A coincis cu o perioadă de ploaie și o creștere a suprafeței de drenaj (Fig. 6.14). În această perioadă s-a observat o dezvoltare intensivă a fitoplanctonului în apele lagunei, din care o proporție semnificativă a fost alge albastru-verde Trichodcsmium (Relevante și colab., 1982). Efectul stimulator al debitului râului asupra dezvoltării fitoplanctonului a fost remarcat și pentru apele de recif din largul Noii Caledonie (Binet, 1984). Datele prezentate indică nefondarea tezei (Parsons et al., 1977) conform căreia comunitățile planctonice din apele tropicale, spre deosebire de apele temperate, se află într-o stare de homeostazie și nu suferă schimbări sezoniere semnificative. Asemenea idei au fost aparent o consecință a lipsei unor cicluri adecvate de observare sezonieră în apele tropicale (Sournia, 1969). Motivul maximelor sezoniere în dezvoltarea fitoplanctonului în apele tropicale de coastă se datorează în primul rând modificărilor circulației apei în zona oceanică adiacentă recifelor, care au o periodicitate anuală. Mai mult, aceste schimbări, chiar și în zona tropicală, au loc cel mai adesea toamna și primăvara. Nu întâmplător maximele sezoniere ale dezvoltării fitoplanctonului sunt observate toamna sau primăvara.
În cursul studierii dezvoltării fitoplanctonului în apele de recif, a fost stabilită prezența unor fluctuații zilnice semnificative în compoziția și abundența comunităților sale, precum și a productivității acestuia (Milliman și Mahnken, 1972; Sournia și Ricard, 1976; Sorokin). , 1979). Astfel de fluctuații apar și în comunitățile de fitoplancton din oceanul deschis. Ele sunt cauzate acolo de periodicitatea zilnică a creșterii fitoplanctonului și de intensificarea consumului de alge pe timp de noapte de către zooplanctonul care migrează din straturile de apă subiacente în zona eufotică. Aceleași motive operează și în apele de coastă din zona recifelor de corali, determinând periodicitate zilnică în dezvoltarea fitoplanctonului. Dar în aceste ape sunt influențate și de ciclul mareelor zilnic. După cum au arătat rezultatele observațiilor zilnice asupra recifelor din apropierea insulelor Namzu, sub influența pășunatului fitoshanctonului care migrează noaptea în coloana de apă de către zooplanctonul recifului, producția sa zilnică dimineața devreme scade cu un ordin de mărime. . Cu toate acestea, până la ora 11 a.m. populația de fitoplancton este în mare măsură restaurată, în special la marginea recifului, unde fitoplanctonul se reproduce intens (Fig. 6.15).
Care sunt motivele creșterii producției de biomasă și fitotancton în apele recifului în comparație cu apele înconjurătoare ale oceanului deschis? Această întrebare este destul de complexă, mai ales având în vedere faptul că o astfel de creștere are loc la concentrații absolute foarte scăzute de nutrienți, nu foarte diferite de cele din oceanul deschis, precum și în condiții de exploatare neobișnuit de intensivă a comunităților de fitoplancton de către filtru. -fauna de hrănire a recifului. Opiniile exprimate pe această temă sunt foarte contradictorii (Sournia, Ricard, 1976; Sournia, 1976b; Salvat 1982). Majoritatea autorilor indică așa-numitul „efect de masă insulă” drept motiv principal. Acest efect constă în îmbogățirea cu substanțe nutritive a apelor din zona de coastă a insulelor datorită interacțiunii masei lor cu fluxul de intrare al curenților oceanici, rezultând ridicarea apelor adânci la suprafață. Efectul îmbogățirii zonei neritice asupra insulelor situate în miezul curenților alizei, cum ar fi, de exemplu, Insulele Marquis (Sournia, 1976c). B. barieră de recif (Gilmartin, Relevante, 1974), Hawaii (Doty, Capurro, 1959), Seychelles (observațiile noastre) apare fără îndoială. Sub influența sa, producția primară în zona neritică a unor astfel de insule este destul de ridicată, ceea ce afectează nivelul de productivitate al apelor recifului. Totuși, chiar și în aceste condiții, producția și biomasa de fitoplancton în apele lagunelor de recif este de obicei mai mare decât în apele zonei neritice adiacente acestora și într-o stare de schimb intensiv cu acestea (Marshall, 1965; Michel, 1969). Sournia, Ricard, 1976, 1976a; Thompson, Golding, 1982). În lumina acestor date, putem concluziona că principalul motiv pentru creșterea productivității fitoplanctonului în apele de recif este eliberarea constantă a elementelor biogene (în primul rând azot, fosfor) în coloana de apă de către heterotrofii de fund și planctonici, care se formează ca un rezultat al proceselor de distrugere a materiei organice care au loc pe recife cu mare forta. În același timp, din cauza ratei foarte ridicate de rotație a nutrienților din coloana de apă a recifelor, nu este întotdeauna posibil să se înregistreze o creștere a concentrațiilor absolute de nutrienți săruri în apele din zona recifului față de oceanul adiacent. ape (Fig. 5.1.).
Fitoplanctonul, prin legarea CO 2 în timpul fotosintezei și formând materie organică, dă naștere tuturor lanțurilor trofice din ocean. Analiza unei varietăți de date privind cantitatea de fitoplancton din diferite zone ale Oceanului Mondial (de la sfârșitul secolului al XIX-lea, calculate din estimările de transparență disponibile și de la începutul anilor 1980, obținute de la distanță de la nave spațiale) arată că biomasa acestuia a scăzut în ultimul secol cu o rată de aproximativ 1% pe an. Cea mai vizibilă scădere a fost observată pentru regiunile oligotrofe centrale ale oceanului. Deși aceste zone se caracterizează printr-o productivitate foarte scăzută, ele ocupă o suprafață imensă și, prin urmare, contribuția lor totală la producția și biomasa fitoplanctonului oceanic este foarte semnificativă. Motivul cel mai probabil pentru scăderea biomasei este creșterea temperaturii stratului de suprafață al oceanului, ceea ce duce la o scădere a adâncimii de amestecare și o reducere a aportului de elemente nutritive minerale din straturile subiacente.
Aproximativ jumătate din producția primară totală a planetei noastre (adică materia organică produsă de plantele verzi și alte organisme fotosintetice) provine din ocean. Principalii producători ai oceanului sunt algele microscopice și cianobacteriile suspendate în straturile superioare ale coloanei de apă (ceea ce se numește colectiv fitoplancton). Studiile cantitative la scară largă ale producției și biomasei de fitoplancton în Oceanul Mondial au început în anii 1960 și 70. Cercetătorii (inclusiv cei de la Institutul de Oceanologie al Academiei de Științe a URSS) s-au bazat apoi pe o metodă bazată pe absorbția de către fitoplancton a izotopului de carbon radioactiv 14 C. Izotopul a fost marcat cu dioxid de carbon CO 2 adăugat la probele de apă cu fitoplancton. ridicat la bordul navei. Ca rezultat al acestor lucrări, au fost construite hărți ale distribuției fitoplanctonului pe tot Oceanul Mondial (vezi, de exemplu: Koblentz-Mishke și colab., 1970). În zonele centrale, mari ale oceanului, biomasa fitoplanctonului și producția acestuia sunt foarte scăzute. Valorile ridicate ale biomasei și producției sunt limitate la zonele de coastă și de upwelling (vezi: Upwelling), unde ape adânci bogate în elemente nutritive minerale ies la suprafață. În primul rând, acestea sunt fosforul și azotul, a căror lipsă limitează creșterea fitoplanctonului în majoritatea apelor oceanice.
O nouă etapă în studiul cantitativ al distribuției fitoplanctonului în Oceanul Mondial a început chiar la sfârșitul anilor 1970, după apariția metodelor îndepărtate (de la sateliți) pentru detectarea apelor de suprafață și determinarea conținutului de clorofilă din acestea. Deși nu mai mult de 10% din fotonii luminii, care se reflectă din apă și poartă informații despre culoarea acesteia, ajung la dispozitivele situate la limita superioară a atmosferei, acest lucru este suficient pentru a calcula cantitatea de clorofilă și, în consecință, biomasa fitoplanctonului (Fig. 1). Valorile biomasei pot fi, de asemenea, utilizate pentru a evalua producția de fitoplancton, care a fost verificată în cursul unor studii speciale, comparând datele satelitare cu rezultatele estimărilor de producție obținute experimental in situ pe vasele de cercetare. Desigur, diferite dispozitive oferă date ușor diferite, dar imaginea de ansamblu a distribuției spațiale a fitoplanctonului și a dinamicii acestuia (sezoniere și interanuale) este foarte detaliată. Este suficient să spunem că dispozitivul Sea WiFS (Sea-viewing Wide Field-of-view Sensor) scanează oceanele întregii lumi în două zile.
O cantitate imensă de date acumulate în ultimii 30 de ani a făcut posibilă identificarea anumitor fluctuații periodice ale biomasei fitoplanctonului, în special asociate cu El Niño, sau, mai precis, cu „Oscilația de Sud” (El Niño-Oscilația de Sud). Analizând aceste materiale, cercetătorii au sugerat existența unor modificări pe termen lung în biomasa fitoplanctonului, dar acestea au fost dificil de detectat din cauza lipsei de date pentru perioada anterioară măsurătorilor satelitului. O încercare de a rezolva cel puțin parțial această problemă a fost făcută recent de specialiștii de la Universitatea Canadian Dalhousie din Halifax (Dalhousie University, Halifax, Nova Scotia). Biomasa fitoplanctonului de acum 50 și chiar 100 de ani poate fi judecată prin estimări ale transparenței, o valoare măsurată în mod regulat în expedițiile de cercetare de la sfârșitul secolului al XIX-lea.

Un instrument de măsurare a transparenței apei, extrem de simplu, dar dovedindu-se foarte util, a fost inventat încă din 1865 de astronomul italian (și în același timp preot) Angelo Secchi, care a fost însărcinat să întocmească o hartă a transparenței Mării Mediterane pentru flota papală. Dispozitivul, numit „disc Secchi” (vezi fig. 2), este un disc de metal alb cu un diametru de 20 sau 30 cm, care este coborât în apă pe o frânghie marcată. Adâncimea la care observatorul încetează să vadă discul este transparența Secchi. Deoarece partea principală a materiei în suspensie care afectează transparența apei este fitoplanctonul, orice modificare a valorii transparenței. de regulă, ele reflectă bine modificările cantității de fitoplancton.

Pe baza estimărilor standardizate de transparență disponibile din 1899 și pe rezultatele unei comparații recente a valorilor de transparență cu concentrațiile de clorofilă, cercetătorii au obținut, în primul rând, o imagine a distribuției biomasei fitoplanctonului în oceane (Fig. 3) și în al doilea rând, modificări ale biomasei fitoplanctonului pe o perioadă de o sută de ani (Fig. 4). În total, au avut la dispoziție rezultatele a peste 455 de mii de măsurători, acoperind perioada 1899-2008. În același timp, datele referitoare direct la zona de coastă (la mai puțin de 1 km de coastă și la adâncimi mai mici de 25 m) nu au fost incluse în mod deliberat în eșantion, deoarece în astfel de locuri influența scurgerii de pe țărm este foarte vizibilă. . Cele mai multe măsurători au fost făcute după 1930 în regiunile nordice ale oceanelor Atlantic și Pacific. Concluzia principală la care ajung autorii este o scădere treptată a biomasei totale de fitoplancton în ultimul secol, cu o rată medie de aproximativ 1% pe an.

Pentru a evalua tendințele locale, întreaga zonă de apă a Oceanului Mondial a fost împărțită într-o grilă cu celule care măsoară 10° × 10°, iar toate valorile au fost calculate ca medii pe celulă. O scădere a biomasei fitoplanctonului a fost observată în 59% dintre celulele pentru care erau disponibile date suficient de fiabile. Majoritatea acestor celule sunt la latitudini mari (mai mult de 60° latitudine). Cu toate acestea, pentru unele zone ale oceanului, a fost observată o creștere a biomasei - în special, în partea de est a Oceanului Pacific, precum și în regiunile de nord și de sud ale Oceanului Indian. Regiunile oligotrofe centrale ale oceanelor au extins efectiv zonele de apă ocupate, iar în aceste zone, în ciuda productivității scăzute, se formează acum aproximativ 75% din producția primară totală a Oceanului Mondial.
Pentru a ne imagina schimbări la nivelul regiunilor mari, întreaga zonă oceanică a fost împărțită în 10 regiuni (Fig. 5): Arctica, Atlanticul de Nord, Ecuatorial și de Sud, părțile de nord și de sud ale Oceanului Indian, Nord, Ecuatoriu. și Pacificul de Sud și Oceanul de Sud. Analiza datelor medii pentru aceste regiuni mari a arătat că o creștere semnificativă a fost observată doar pentru partea de sud a Oceanului Indian și o creștere nesigură din punct de vedere statistic pentru partea de nord a Oceanului Indian. Pentru toate celelalte regiuni, a fost observată o reducere semnificativă a biomasei fitoplanctonului.

Discută despre posibilele motive pentru modificările observate, autorii acordă atenție în primul rând creșterii temperaturii stratului de suprafață al coloanei de apă. A acoperit aproape tot oceanul și a dus la scăderea grosimii stratului mixt. În consecință, afluxul de elemente nutritive minerale (în primul rând fosfați și nitrați) din straturile subiacente este redus. Cu toate acestea, autorii admit că o astfel de explicație nu este potrivită pentru latitudini mari. Acolo, încălzirea stratului superior ar trebui să crească, mai degrabă decât să scadă, producția de fitoplancton și biomasa. În mod clar, mecanismele care determină schimbări la scară largă în biomasa fitoplanctonului necesită studii suplimentare.
„Academia Rusă de Științe
UDC 574.583(28):o81 +574.55:58.035
PYRINL Inna Lopshovna
CA FACTOR DE PRODUCTIVITATE A FITOPLANCTONULUI ÎN CORPELE DE APĂ INTERIORĂ
03.00.16 - ecologie
D i s e r i a c i n ||, h concurs pentru un om de știință cu diplomă în științe biologice sub forma unui raport științific
Sankt Petersburg 1995
Lucrarea a fost efectuată la Institutul de Biologie al Apelor Interioare, numit după. II. D. Papashsha RAS.
Adversari oficiali:
Doctor în științe biologice Lavrentyeva G. M. Doctor în științe biologice Bulyon V. V. Doctor în științe biologice Raspopov I. M.
Instituție lider: Institutul de Ecologie al Bazinului Volga RAS „.....
Apărarea va avea loc „x. /" aprilie 1995 la "ora. la o ședință a consiliului de specialitate D 200.10.01 la Institutul de Știință a Lacului al Academiei Ruse de Științe (198199, Sankt Petersburg, Sevastyanova St., 9).
Dizertația sub formă de raport științific poate fi găsită în biblioteca Institutului de Știință a Lacului al Academiei Ruse de Științe.
Secretar stiintific al consiliului de specialitate
Candidat la Științe Biologice
M. A. Belova
Introducere
Relevanța cercetării
În legătură cu deteriorarea stării ecosistemelor acvatice, cercetarea proceselor care influențează formarea calității apelor naturale și, mai ales, fotosinteza planctonului, datorită căreia rezervorul este completat cu materie organică primară și oxigen. devenind din ce în ce mai important. Cel mai important factor pentru acest proces, care are loc într-un mediu acvatic, este lumina. Acest lucru se datorează accesului limitat la plancton al luminii solare, a cărui parte principală este absorbită și împrăștiată de apă și substanțe conținute în neii și nu ajunge la celulele fotosensibilizante. În plus, compoziția spectrală a radiațiilor penetrante se modifică în apă - razele roșii și albastre, utilizate în principal în fotosinteză, sunt cel mai puternic reținute, în timp ce razele verzi sunt transmise mai complet. Ca urmare, fitocenozele planctonice, având un avantaj față de cele terestre în ceea ce privește compușii carbonatați și apei necesari pentru fotosinteză, elementele de nutriție minerală și o temperatură ambientală destul de stabilă, sunt dezavantajate din partea sursei de energie. Și dacă pe suprafața Pământului, care primește relativ puțină lumină solară slăbită de aer, plantele nu duc lipsă de energie luminoasă, iar fotosinteza lor este limitată de alți factori, atunci sub apă acest proces este cel mai limitat de lumină.
În teoria productivității fotosintetice a vegetației, dezvoltată de cercetătorii fitocenozelor terestre, se acordă o mare importanță energiei radiației solare ca factor în formarea culturilor (Nichiporovich 1956, 1908). Hidrobiologii, atunci când evaluează producția primară a ecosistemelor acvatice, apelează relativ rar la acest factor, mai ales atunci când lucrează pe corpurile de apă dulce, care, însă, au cel mai mare nevoie de acest gen de cercetări din cauza diversității mari din punct de vedere hidrooptic. Prin urmare) studii speciale ale factorului de lumină al fotosintezei subacvatice, începute pe lacuri în anii 30 (Schomer, J934; Schomer, Juday, ¡935: Manning, Juday, 1941) și preluate în anii 50-80 (Talling, ¡ 957, 1971, 1982; Tilzer, Schwarz, 1976; Tilzer, ¡984; Ganf, 1975; Jewson, 1976, 1977; Kirk, 1977, 1979 - citat în; Kirk, 1983; Kirk, 1983; Kirk, 1983; Kirk, 1983; 1983; Megard și colab. Mai mult, dacă îl comparăm cu studiul factorului biogen al productivității fitoplanctonului, spre care studiile limnologice din ultima perioadă au deviat din cauza problemei eutrofizării corpurilor de apă.
Studiul luminii ca factor în fotosinteza subacvatică este indisolubil legat de studiul pigmenților fitoplanctonului care captează energia luminii. Principala este clorofila „a”, fiind o substanță specifică tuturor plantelor și regiunilor de fotosinteză.
oferind proprietăți spectrale unice, face posibilă cuantificarea biomasei algelor fără a le separa de restul planctonului. În calitate de participant direct la procesul de fotosinteză, poate servi simultan ca un indicator al activității de asimilare a fitoplanctonului. Alți pigmenți, dintre care mulți sunt unici pentru alge, sunt importanți pentru înțelegerea ecologiei fotosintezei subacvatice în condiții de lumină care variază în adâncime.
Ideea utilizării clorofilei în determinarea biomasei planctonului fotosintetic (Vinberg, 1954, 1960) s-a dovedit a fi atât de fructuoasă încât a servit drept bază pentru dezvoltarea unor cercetări ample privind studiul acestui pigment în diferite tipuri de apă. organisme cu dezvoltarea unor metode speciale de analiză, inclusiv direct în apă în regim continuu și de la distanță. Definiția clorofilei a fost inclusă în majoritatea lucrărilor hidrobiologice, unde este considerată un indicator al productivității și calității apelor naturale și a devenit parte integrantă a „monitorizării de mediu a corpurilor de apă. Creșterea interesului pentru acest pigment. în hidrobiologia modernă s-a dovedit a fi cu nu mai puțin de aproximativ 100 de ani în urmă, când a fost descoperit rolul în fotosinteza plantelor.Cu toate acestea, studiile asupra proprietăților specifice ale clorofilei, care asigură absorbția energiei solare de către plancton, precum și rolul de alți pigmenți din fotosinteză sub lumina subacvatică, în special în corpurile de apă dulce, rămân puțini (Tilzer, 1983; Ganf, et al., 1991) -.
Între timp, parametrii care caracterizează relația dintre fotosinteza algelor planctonice și energia radiației solare și a clorofilei stau la baza metodelor de calcul larg răspândite pentru determinarea și modelarea matematică a producției primare de fitoplancton. Este important să cunoaștem valorile acestor parametri care sunt cele mai adecvate unui anumit mediu natural. Oceanologii efectuează lucrări foarte aprofundate în această direcție (Platt și colab., 1980, 1990; și alții), inclusiv pe baza experimentelor naturale (Koblenz-Mishke, 1980; Koblenz-Mishke și colab., 1985; 1987). În corpurile de apă de apă dulce, astfel de cercetări sunt mai puțin dezvoltate, iar parametrii necesari modelării fotosintezei subacvatice se regăsesc în principal teoretic sau din surse din literatură (StraSkraba, Gnauck, 1985).
Scopul și obiectivele cercetării
Lanțul principal a constat în identificarea relației dintre condițiile de lumină subacvatică și conținutul de pigmenți de fitoplancton în timpul fotosintezei și formarea producției primare în ecosistemele de apă dulce.
În acest scop, au fost stabilite următoarele sarcini specifice: (1) să studieze modelele de intrare și pătrundere a energiei radiației solare din spectrul general și din regiunea activă fotosintetic în corpuri de apă dulce optic diferite, ținând cont de influența soluției.
substanțe colorate libere, suspensie generală și celule de fitoplancton; (2) se determină conținutul de clorofilă ca indicator al biomasei de fotosinteză, raportul acesteia cu alți pigmenți fitoplanctonici, se studiază modelele modificărilor lor spațiale, sezoniere și interanuale, se evaluează diferențele asociate stării trofice a corpurilor de apă studiate; (3) să determine nivelul producției primare de fitoplancton și eficiența utilizării acestora a energiei solare în comparație cu condițiile de lumină și cantitatea de clorofilă; (4) evaluarea rolului energiei solare care intră pe Pământ în fluctuațiile interanuale ale productivității fitoplanctonului și eutrofizarea rezervorului; (5) să studieze dependența de lumină a fotosintezei fitoplanctonului cu un set divers de pigmenți într-un mediu natural și posibilitatea utilizării acestora pentru a modela producția primară.
Dispoziții protejate
I. Cantitatea de energie luminoasă disponibilă pentru fitoplancton și clorofila care îl captează determină nivelul producției primare a corpurilor de apă. 2. Cu arhitectura relativ simplă a fitopoiezei planctonice, modelele de penetrare a luminii solare intensificate în timpul procesului de fotosinteză și într-un rezervor sunt destul de ușor descrise matematic. 3. Aceasta deschide perspective pentru dezvoltarea metodelor de calcul pentru evaluarea producției primare de fitoplancton pe baza caracteristicilor măsurate instrumental.
Cercetare științifică
1. Folosind echipamentul original, primele măsurători ale energiei radiațiilor active fotosintetic subacvatice au fost efectuate într-o zonă mare de rezervoare de apă dulce (rezervoare Volga, lacurile Pleshcheezo, Onega). Au fost studiate modelele de penetrare a acestuia în diferite tipuri optic de ape, sub aspect latitudinal și sezonier, inclusiv în perioada sub-gheață, cu evaluarea ponderii în spectrul solar total. a fost relevat cel descris de legea clasică Bouguer, caracteristic fluxului complex spectral al razelor solare în apele naturale și s-a găsit un parametru care să compenseze această abatere în mod matematic. Contribuția fitoplanctonului în raport cu alte materii suspendate în ecologizarea s-a determinat radiaţia penetrantă.
2. Pentru prima dată, conținutul de pigmenți de fitoplancton a fost determinat în rezervoarele și lacurile din bazinul Volga, măsurile Ladoga și Onega, lacurile tundra Kharbey, râul. Yenisei cu o evaluare a proporției de ch.chozophylla "a" în cantitatea de derivați ai săi și cu alte clorofile, precum și raportul dintre cantitatea totală de pigmenți verzi și galbeni." Modelele distribuției lor spațiale, sezoniere și au fost studiate variabilitatea pe termen lung.Primele date privind specific
4. În timpul cercetărilor din rezervorul Rybinsk, a fost obținută una dintre cele mai lungi serii de observații pe termen lung din lume (27 de ani) ale conținutului de clorofilă. Este prezentată legătura dintre fluctuațiile sale interanuale și caracteristicile sinoptice ale diferiților ani. S-a identificat o tendință de creștere a nivelului concentrațiilor de pigment, indicând eutrofizarea rezervorului și rolul energiei radiației solare care intră pe Pământ în acest proces.
5. O serie de rezervoare (anumite zone din V. Olga înainte de reglementare, lacul Ivankovskoe, lacurile Dvina de Nord și Kharbey, Lacul Pleshcheyevo, Lacul Ladoga) au fost studiate pentru prima dată în legătură cu producția primară de fitoplancton.
6. În rezervoarele studiate au fost evaluate pentru prima dată eficiența utilizării energiei radiației solare de către fitoplancton și relația acesteia cu condițiile de lumină și conținutul de clorofilă.
7. Pe baza experimentelor originale în cadru natural, s-au obținut noi date despre dependența de lumină a fotosintezei fitoplanctonului divers din punct de vedere ecologic și taxonomic. Se oferă o reprezentare analitică a acestei dependențe.
8. Au fost realizate mai multe dezvoltări metodologice noi care și-au găsit aplicație în studiul indicatorilor producției primare de fitoplancton: (1) s-au proiectat echipamente pentru măsurarea radiației active fotosintetice subacvatice și s-a propus o metodă simplă de calibrare a acesteia în unități de iradiere. bazat pe metode actinometrice; (2) analiza spectrofotometrică a pigmenților fitoplanctonului din extractul total a fost introdusă în practica cercetării hidrobiologice din țară; (3) a fost elaborată o metodă de calcul pentru determinarea producției primare de fitoplancton pe baza intensității aportului. radiația solară rezervor și dependența de lumină a fotosintezei subacvatice; (4) procedura de calcul a caracteristicilor medii fitoplanctonului pentru un rezervor și perioadă sezonieră a fost îmbunătățită; (5) au fost dezvoltați algoritmi originali pentru calcularea erorilor de măsurare și medierea indicatorilor de productivitate fitoplanctonului folosind metoda de evaluare a erorilor indirecte, comună în disciplinele exacte;
Valoare practică
Întrucât activitatea fotosintetică a fitoplanctonului este asociată cu îmbogățirea corpurilor de apă cu materie organică și oxigen, studiile efectuate în cadrul acestei lucrări au făcut parte dintr-o serie de proiecte care vizează evaluarea calității apelor naturale în starea lor naturală și sub diferite tipuri de influență antropică. Aceasta este (1) lucrare propusă de Comitetul guvernamental pentru știință și tehnologie (GKNT) pentru a identifica rolul apelor de mică adâncime în modelarea calității apei din Rybinsk și Ivankovsky
rezervoare (1971 - 1973, raport în 1973), conform evaluării stării ecologice a lacului de acumulare Ivankovo, inclusiv după punerea în funcțiune a centralei electrice din districtul de stat Konakovo (1970 - 1974, raport în 1975) și rezervoare a Volga- sistemul baltic, inclusiv lacul de acumulare Rybinsk, în legătură cu problema redistribuirii debitului râului (1976 - 1985, raport în 1980 și 1985); (2) experimente aerospațiale în rezervorul Rybinsk cu scopul de a dezvolta metode la distanță de monitorizare a productivității rezervorului pe bază de pigmenți de fitoplancton (1986 - 1990, raport în 1990); (3) lucrările efectuate pe baza instrucțiunilor organelor administrative regionale și organizațiilor practice, cum ar fi monitorizarea mediului lacului. Pleshcheev (1986 - 1992, rapoarte în 1986, 1990 și 1992); evaluarea consecințelor lucrărilor privind îndepărtarea amestecului de nisip și pietriș din sol pentru biota rezervorului - Rezervorul Kuibyshev (1990 - 1991, raport în 1991); (4) lucrează în cadrul unui acord de cooperare creativă cu Universitatea de Stat din Krasnoyarsk privind studiul clorofilei în corpurile de apă, având ca scop diseminarea acestei metode în practica monitorizării mediului în corpurile de apă (1986 -1987, raport în 1987); (5) ) lucrează în cadrul programului de mediu „Omul și Bnoefer” (proiectul nr. 5) pentru identificarea factorilor care influențează nivelul producției primare în rezervoare (1981 - 1990 cu rapoarte anuale, precum și cele consolidate în 1986, 1988 și 1991). ).
Aprobarea lucrării
Rezultatele și principalele prevederi ale lucrării au fost prezentate la prima întâlnire privind producția științifică a rezervoarelor (Minsk, 1960: la reuniunea întregii Uniuni privind problema rolului algelor verzi în alte rezervoare ale URSS (Korok, 1960). *: la 1. P, Sh. VI Întrunirile limnologice ale întregii uniuni asupra orbului și energiei și rezervoarelor lacurilor (Lisgvenichnoye-on-Bankale, |"64, 196"-). 1973, 1985); la I (Moscova, !% 5] , I (Kshshshev, 197I.). V !ol! din i. 1986) a.ezlnkh VGBO: la 1 (Tolyatti. 1968) si II (Kor. ¡974 ¡.; conferinte si ¡studii asupra rezervoarelor Volga). bazin; despre utilizarea comună integrată a rezervoarelor (Kiev, 1997); la simpozionul nervos privind hidrologia și biologia rezervoarelor încălzite sub centrale termice (Eorok, 197!): GTA TU (Kiev, 1972). ) și XI! (Lnstvenichnoye-ia-Bankale, 1984) Întruniri ale întregii uniuni" pe actinometrie; la cel de-al II-lea Simpozion al întregii uniuni privind problema corpurilor de apă eptrofine (Zvesh!gorod, 1977); la a II-a Conferință a întregii uniuni „Probleme ale ecologiei din regiunea Baikal” (Irkutsk, 1982); la o întâlnire privind problemele productivității biologice, utilizării raționale și protecției corpurilor de apă din regiunea Vologda (Vologda, 1978); la Reuniunea științifică a întregii uniuni „Resurse naturale ale lacurilor mari ale URSS” (Leningrad, 1982); la o întâlnire regională privind problemele protecţiei şi utilizării raţionale a intern
apele regionale din Centrul și Nordul Câmpiei Ruse (Iaroslavl, 1984); la I și I Întâlniri internaționale privind problemele sondajului aerospațial al apelor interioare (Leningrad, 1987, 1988); la Şcoala-seminar All-Union „Metode cantitative în hidrobiologie” (Borok, 1988); la I Conferința Internațională Vereshchagin Baikal (Listvenichnoe-on-Baikal, 1989); la întâlniri metodologice privind studiul producției primare de plancton în corpurile de apă interioară (Borok, 1989) și evaluarea productivității fitoplanctonului (Irkutsk, 1992); la o întâlnire a Asociației Biologice de Apă Dulci din Marea Britanie (Windermere, 1990); la conferința științifică panrusă dedicată aniversării a 300 de ani a flotei ruse (Pereslavl-Zalessky, 1992); la conferința privind problemele de mediu în studiul râurilor mari (Tolyatti, 1993); la o întâlnire privind observațiile hidrobiologice pe termen lung asupra apelor interioare (Sankt Petersburg, 1994); precum și la seminarii și întâlniri științifice la locul de muncă la Institutul de Biologie a Apelor Interioare al Academiei Ruse de Științe.
Publicaţii
Rezultatele cercetării sunt prezentate în 15 secțiuni din 10 monografii colective și în 65 de reviste și alte articole. Înainte de a-și susține teza de doctorat, au fost publicate 10 articole pe această temă.
I. Materiale şi obiecte de cercetare
Lucrarea se bazează pe rezultatele studiilor privind condițiile de lumină, conținutul de pigment și intensitatea fotosintezei fitoplanctonului în rezervoarele Volga și rezervoarele adiacente, în lacurile Kharbey din tundra, efectuate pe parcursul întregului sezon de vegetație (rezervor Ivankovskoye - 1958, 1970 - 1971, 1973 - 1974; lacul de acumulare Rybinsk - 1958, 1969-1973; lacul de acumulare Kuibyshev - 1958; lacul Beloe 1976-1977; lacul Pleshcheyevo - 1983-1985; studiile de rută în anumite perioade ale râurilor sau în anumite perioade Kharbey -1985. 57 , 1960; rezervoare ale căilor navigabile Volga-Baltice și Dvina de Nord, inclusiv lacurile Ladoga și Onega - 1973; rezervoare Sheksna și Volga Superioară - 1979). Observațiile pigmenților de fitoplancton din zona de conservare Ivankovsky au continuat în 1977-1978, în lac. Pleshcheyevo - până în 1991, în lacul de acumulare Rybinsk s-au mutat în cercetare continuă pe termen lung, care este în desfășurare până în prezent. Studii detaliate ale pigmenților fitoplanctonului au fost efectuate la gistatoriul lacului Onega (1967-1968) și al râului. Yenisei (1984-1985). În rezervorul Rybinsk, modificările la scară mică ale conținutului de pigment au fost studiate ca parte a unui experiment aerospațial pentru a dezvolta o tehnică de detecție optică de la distanță a corpurilor de apă.
în zona apei şi în timp (1986-1988). În rezervoarele Rybinsk (1971-1972) și Ivankovsky (1973-1974), au fost efectuate o serie de lucrări în zona de mică adâncime pentru a evalua rolul acesteia în îmbogățirea întregului rezervor cu materie organică primară. În rezervorul și lacul Rybinsk. Pleshcheyevo a efectuat observații de iarnă ale dezvoltării fitoplanctonului în condițiile regimului de lumină al perioadei subglaciare. În timpul expedițiilor de rută către rezervoarele Volga și Sheksninsky (i960, 1979), în rezervorul Rybinsk (1970-1971, 1987), în lacurile Onega (1968) și Pleshcheyevo (1983-1984), au fost efectuate lucrări speciale privind studiul radiației solare. pătrunzând în dependențele de apă și lumină ale fotosintezei subacvatice. Activitatea de fotosinteză a fost studiată în rezervorul Ivankovo. planctonul în condiții de temperatură ridicată (1970-1971), creat sub influența apei uzate dintr-o centrală termică, în rezervorul Kuibyshev - efectul asupra fitoplanctonului al suspensiilor minerale care intră în rezervor în timpul dragării și altor lucrări. legate de îndepărtarea solului (1990-1991).
Unele materiale au fost obținute împreună cu angajații Institutului de Biologie a Apelor Interioare (J1.B. Morokhovets, O.I. Feoktistova, N.P. Mokeeva, A.L. Ilyinsky, V.A. Elizarova, E.I. Naumova, V.G. Devyatkin, L.E. Sigareva, E.L.Kato, N.M.G. Korneva, V.L. Sklyarenko, A.N. Dzyuban, E.G. Dobrynin, M.M. Smetanin ) și alte instituții științifice (V.A. Rutkovskaya, I.I. Nikolaev, M.V. Getsen, T.I. Letanskaya, I.S. Trifonova, T.N. au fost scrise sau au fost folosite în această lucrare articolele lor independente care conțin datele necesare. T.P. a participat întotdeauna la colectarea și prelucrarea materialelor de mulți ani. Zaiknna este asistent principal de laborator la laboratorul de algoologie al Institutului de Biologie a Apelor Interioare. Tuturor acestora, precum și personalului centrului de calculatoare și atelierelor experimentale, personalului tehnic al Institutului de Biologie a Apelor Interioare, studenților din universitățile Moyek, Vsky, Sankt Petersburg, Nijni Novgorod, Yaroslavl, Perm și Kazahstan, care au avut stagii de practică la Institut, autorul își exprimă profundă recunoștință și recunoștință pentru ajutorul acordat în realizarea cercetărilor.
II. Metode de cercetare
Studierea caracteristicilor modului de lumină
Studiul condițiilor regimului luminos ca factor de producție primară a fitoplanctonului este asociat cu dificultăți metodologice semnificative din cauza faptului că energia radiantă a Soarelui care trece prin se modifică nu numai cantitativ, ci și calitativ. Compoziția spectrală și unghiulară
Caracteristicile radiațiilor penetrante, intensitatea radiațiilor care intră în apă se modifică continuu în funcție de înălțimea Soarelui deasupra orizontului și de condițiile norului. În mod ideal, este necesar un dispozitiv care să poată înregistra atât radiația care variază adâncimea, cât și radiația fotosintetic activă însumată în timp - PAR (k = 380-710 nm), exprimată în unități de iradiere, deoarece astfel de dispozitive nu existau (Raport... .. ., 1965, 1974). Dispozitivul constă dintr-un set de senzori care captează radiația din emisferă - iradierea (supusă legii cosinusului), dintre care unul este sensibil în intervalul de lungimi de undă care acoperă PAR (380-800 nm), ceilalți - în secțiuni mai înguste ale acestui regiunea spectrului (480-800, 600-800, 680-800 nm). Sensibilitatea spectrală a senzorilor se realizează prin combinarea unei fotocelule în vid TsV-3 cu filtre de lumină SZS-14 + BS-8 - care acoperă întreaga gamă de lungimi de undă de 380-800 nm și SZS-14 + ZhS-17, SZS-14 + KS-10, SZS-14 + KS-19 - pentru restul părților sale care se înclină treptat. Dispozitivul de înregistrare însumează automat energia radiației în timp.
Calibrarea dispozitivului în unități de iradiere, dezvoltată pe baza metodelor actinometrului (Berezkin, 1932), se realizează în funcție de radiația solară directă, măsurată cu un actinometru cu filtre de lumină adecvate. Razele directe sunt separate de fluxul total de radiație care ajunge la senzor folosind un tub montat pe acesta, care este similar ca design cu tubul actinometrului (Pyrina, 1965, 1993).
Mulți ani de experiență în utilizarea unui fotointegrator subacvatic, inclusiv în comparație cu dispozitivele actinometrice standard, au demonstrat fiabilitatea funcționării acestuia în condiții de câmp cu o precizie suficient de mare a rezultatelor măsurătorilor atât a fluxului PAR integral, cât și a secțiunilor înguste ale acestei regiuni spectrale. . Primele mostre făcute acum aproximativ 30 de ani încă funcționează. Mai mult decât atât, dispozitivele seriale pentru măsurători subacvatice ale rețelelor fază, cum ar fi Li-Cor și QSP (SUA), QSM (Suedia), existente în prezent (Jewson și colab., 1984), precum și modele unice în țara noastră (Semenchenko și colab. , 1971; Cehia, 1987), sunt încă inaccesibile.
Aproape toate experimentele de determinare a producției primare de fitoplancton au fost măsurate prin măsurători ale energiei integrate peste spectrul PAR care intră în rezervor; în acest scop, a fost instalat un senzor fotointegrator pe partea superioară a suprastructurii navei sau pe o cotă deschisă către Soare pe mal, înregistrând afluxul de radiații în timpul de expunere.
La studierea condițiilor regimului de lumină într-un rezervor, măsurătorile au fost efectuate în mai multe părți ale spectrului PAR, folosind întregul set de senzori fotointegratori subacvatici, diferiți în sensibilitatea spectrală. Senzorii au fost lansați în rezervor suspendați pe un flotor, împreună cu care au fost îndepărtați de vas la 10-15 m, mai rar pe un troliu echipat cu prelungire alungită în direcția Soarelui. Unele dintre aceste măsurători au fost însoțite de observații ale pătrunderii radiației solare în spectrul general, care au fost efectuate în 1960 de V.A.Rutkovskaya (1962, 1965) și în 1979 de T.N. Pokatilova (1984, ¡993). Onn a folosit piranometrul Yu.D. Yashpievsky (1957), adaptat pentru măsurători subacvatice, care a fost scufundat într-un rezervor cu troliu cu îndepărtare. Toate măsurătorile radiației penetrante au fost efectuate la 10-12 orizonturi până la o adâncime, până când sensibilitatea de prag a instrumentelor a făcut posibilă obținerea unor citiri fiabile.
În acele cazuri în care măsurătorile radiației solare au fost folosite pentru a efectua experimente privind dependența de lumină a fitoplanctonului, pe dispozitiv au fost montate baloane cu probe de testare, iar o astfel de instalație a fost expusă în rezervor pe toată durata expunerii. Aceste instalații au fost dotate cu numărul de orizonturi pentru determinarea fotosintezei, planificat în funcție de transparența apei. În acest fel, a fost posibilă înregistrarea completă a energiei de radiație care intră în probă și obținerea datelor care erau cele mai adecvate dependențelor de lumină ale fotosintezei planctonului natural.
Dacă este necesar, aveți date despre condițiile de lumină subacvatică
pentru un număr mare de stații au recurs la o metodă de calcul pentru determinarea radiației penetrante pe baza sosirii acesteia la momentul t; Boi de transparență Coc Sekhki, folosind F.E. propus, Api c D.I. Tolstyakov (1969) formula sau, mai târziu, un pari chit rafinat (Pyrina, 1989).
izanmotrans.chod din energia radiației de intrare a spectrului general:: (l. = 380-710 im) sau înregistrat de un fotointegrator (l = 3íW-300 i, i) și invers a fost efectuat folosind coeficienți empirici (Pyrnna , 1985), selectat de pavnenmoesh sau de relații despre personalitate, iar pe vreme senină - tot despre înălțimea Soarelui.
Datele despre fitoplancton ingmengs - conținutul total de clorofile, datând din 1958 (Pyrpna, I960), au fost obținute prin metoda fotometrică (Vinberg, Sivko, 1953) cu calibrare folosind un extract din culturi de diatomee și alge albastre-verzi, unde concentrația inițială de clorofile a fost măsurată spectrofotometric pe baza coeficientului specific de extincție 95 l/g cm (Koski Smith, 1948).Din 1960, se folosește o metodă spectrofotometrică pentru determinarea formelor individuale de clorofilă și carotnoide în extractul total (Richards, Thompson). , 1952). Această metodă este
valoroasă apoi pentru analizele pigmenților fitoplanctonici pentru prima dată în țară (Pyrna, 1963) și concomitent cu cercetări străine avansate în această direcție (Humphrey, 1963; Tailing, Driver, 1963), apoi s-au răspândit în studiul pigmenților fitoplanctonici și , după unele clarificări (Parsons, Strickland, 1963; SCOR-UNESfCO, 1966; Jeffrey, Humphrey, 1975), a fost recomandată ca standard (Lorenzen, Jeffrey, 1980; Marker et al., 1982; GOST, 1990).
Concentrațiile de clorofilă au fost calculate în 1960 folosind formulele lui Richards și Thompson (1952), în 1967-1976. - SCOR-UNESCO (1966), în alți ani - Jeffrey și Humphrey (1975). Concentrațiile de carotenoide au fost calculate în primul an folosind formulele lui Richards și Thompson (1952), apoi Parsons și Strickland (1963) pentru ziua de plancton de diatomee care a predominat în rezervoarele studiate. Concentrațiile de feopigmenți și, minus acestea, clorofila a pură au fost calculate folosind formulele lui Lorenzen (Lorenzen, 1967).
După cum se știe, formulele lui Richards și Thompson (1952) au folosit coeficienți de extincție specifici subestimați ai clorofilei a și b, precum și valorile sale condiționate pentru clorofila c și carotenoide și, prin urmare, concentrațiile acestora diferă de cele obținute ulterior. Pentru clorofila „a” acestea sunt supraestimate cu 25% față de cele calculate folosind alte formule bazate pe coeficienți de extincție mai mari ai acestui pigment (88-92 l/g.cm). Datele privind concentrațiile de clorofilă „b” și „c” s-au schimbat mai mult pe măsură ce coeficienții de expansiune și formulele pentru calculul lor au fost rafinați - cu 150-200%. Rezultatele calculelor folosind diferite formule pentru carotenoizi diferă și mai mult (de până la 2,5 ori); datorită diversității compoziției lor în plancton natural, este aproape imposibil să se selecteze valori adecvate ale coeficienților de extincție pentru calcularea concentrațiilor acestor pigmenți. în extractul total. Prin urmare, mai târziu, proporția carotenoidelor față de clorofiați a fost judecată după raportul disparițiilor în regiunea cu cea mai mare contribuție a acestora la absorbția luminii de către extractul - Eva / (Pyrina, Sigareva, 1976), propus în aceste scopuri încă la sfârșitul anilor '50. (Burkholder şi colab., 1959). Luând în considerare abaterile observate în rezultatele calculelor din diferiți ani, compararea datelor privind nivelul pigmenților fitoplanctonului a fost efectuată folosind clorofila „a”, ca cea mai precisă determinată. În același timp, a fost introdusă o corecție de 0,75 la datele timpurii obținute folosind formulele Richards și Thompson (Pyrina și Elizarova, 1975).
În procesul de lucru asupra metodei de determinare a pigmenților de fitoplancton, a fost testată o metodă neextractivă de măsurare a clorofilei direct în celulele de alge colectate pe filtre cu membrană, după ce acestea au fost clarificate cu ulei de imersie (Yentsch, 1957). Metoda a atras atenția pentru simplitatea ei și a arătat rezultate satisfăcătoare în analiza planctonului de lac (Vinberg și colab., 1961 Totuși, s-a dovedit a fi inacceptabilă pentru determinarea
deficit de clorofilă în rezervoare caracterizat printr-un conținut ridicat de detritus și substanțe minerale în suspensie (Pyrina și Mokeeva, 1966).
3. Determinarea producţiei primare de fitoplancton
În studiile staționare în rezervoare mici de lac (Lacul Plesheevo, Lacurile Kharbey), au fost efectuate experimente clasice „in situ” (Vinberg, 1934) cu eșantionare pentru a măsura fotosinteza la adâncimea expunerii lor - 5-7 orizonturi din zona eufotică a rezervorul. Pe rezervoarele cascadei Volga și rezervoarele adiacente, unde s-a desfășurat lucrări de pe o navă de expediție, a fost utilizată o metodă de calcul pentru a determina producția primară cu expunerea eșantionului pentru a măsura valorile inițiale ale fotosintezei într-un incubator de punte care simulează condițiile de stratul superficial de apă. În același timp, în prima etapă a cercetării, au fost efectuate experimente conform schemei modificate a lui Yu.I. Sorokin (1958), care prevede măsurarea intensității fotosintezei în probe de mai multe adâncimi ale zonei fotonice a rezervor cu o legătură empirică la valorile obținute pentru atenuarea radiației solare penetrante (Pyrina, 1959(a)), 1966). Ulterior, s-au limitat la determinarea valorii maxime a fotosintezei de-a lungul profilului vertical din incubator (Amshs) pe baza probei medii sau totale pentru zona eufotică, iar scăderea acesteia în adâncime a fost evaluată pe baza pătrunderii radiațiilor (Pyrina, 1979). Energia radiației la adâncimile studiate a fost de obicei găsită în astfel de experimente folosind metode computaționale (Are și Tolstyakov, 1969; Pyrina, 1989, 1993). Dacă a fost imposibil să se determine intensitatea fotosintezei, a fost utilizată metoda clorofilei cu o estimare a Alm din numărul de asimilare, care a fost selectat din valorile stabilite anterior, în conformitate cu condițiile specifice ale rezervorului.
Experimentele pentru studiul dependențelor de lumină ale fotosintezei fotosintezei au fost efectuate într-un mediu natural cu expunere la mai multe adâncimi a zonei efotice a unui rezervor omogen”; probe de fund prelevate de la suprafață sau, în cazul stratificării pronunțate, complet în interiorul epilnmnonului. De regulă, la aceleași adâncimi pe tot parcursul expunerii, au funcționat senzori de fogointegrator, adesea li se atașează baloane cu probe de testat (Pyrina, 1967, 1974).În unele experimente, împreună cu PAR înregistrat de fogointegrator la același adâncimi, s-au efectuat măsurători urgente ale energiei radiațiilor din spectrul general cu ajutorul unui piranometru subacvatic și s-au calculat sume în timpul de expunere, ca urmare s-au obținut o serie de curbe luminoase de fotosinteză, construite în raport cu radiația solară totală. (Pyrina, Rutkovskaya. 1976) Dacă ar fi imposibil să păstrați instrumentele lângă chks pentru o lungă perioadă de timp: măsurători unice ale iradierii subacvatice au fost făcute cu mostre blocate, de obicei la amiază, la baza muntelui? și energia PAR care vine la suprafața rezervorului. continuu rs
colectat pe mal sau la bordul unei nave, s-a determinat sosirea acestuia la adancimile studiate in timpul expunerii.
Intensitatea fotosintezei a fost evaluată prin oxigen în timpul expunerii zilnice, care în experimentele staționare „in situ” au început seara, în altele - pe măsură ce probele au fost prelevate în stații, cu respectarea recomandărilor de bază disponibile în literatura de specialitate cu privire la această metodă (Alekinidr ., 1973; Vollen Weider şi colab. (1974; Pyrina, 1975, 1993).
4. Prelucrarea matematică a rezultatelor cercetării
Întrucât, pe baza rezultatelor cercetării, nivelurile de productivitate fitoplanctonului au fost comparate în diferite rezervoare și în diferiți ani, când este important să avem confirmarea fiabilității diferențelor emergente, s-a acordat o atenție deosebită aprecierii valorilor medii din datele obținute și erori în reprezentativitatea acestora (Pyrina, Smetanin, 1982, 1993; Pyrina, Smetanii, Smetanina, 1993).
Determinarea valorilor medii pentru un rezervor este complicată de faptul că acestea se referă la materiale de origine diferită - măsurători repetate într-un singur eșantion, probe de la diferite adâncimi la stații individuale, diferite stații și secțiuni ale rezervorului și diferite perioade ale sezonul de creștere. Prin urmare, medierea a fost efectuată în mai multe etape, la unele dintre ele, când s-a făcut media datelor pentru mase de apă de diferite volume sau pentru diferite intervale de timp, valorile medii au fost calculate ca medii ponderate.
Procedura de mediere a datelor pe suprafața de apă a unui rezervor a inclus calcularea mediei aritmetice pe baza rezultatelor măsurătorilor repetate dintr-o probă, apoi pe baza datelor de eșantion pentru aceleași adâncimi ale diferitelor stații (sau straturi de apă), apoi un medie aritmetică ponderată pe baza datelor din straturile individuale pentru fiecare secțiune și, în final, rezervorul în ansamblu.Ultimele două valori au fost calculate luând în considerare volumele de straturi și respectiv secțiuni ale rezervorului, estimate prin nivelul acestuia la timpul de observare În absența datelor batimetrice necesare pentru astfel de evaluări, a fost permisă echivalarea bazinului rezervorului cu un corp cu pereți verticali și în locul volumelor secțiunilor sale, aria lor a fost introdusă în calcule și în locul volumelor de straturi de apă, grosimea acestora a fost introdusă în calcule.
Obținerea datelor medii pentru sezonul de vegetație s-a redus la calcularea mediilor aritmetice pentru intervalele de timp dintre perioadele individuale de observare, apoi a unei medii aritmetice ponderate pentru întreaga perioadă, ținând cont de numărul de zile din fiecare interval. Sezonul de vegetație din rezervoarele studiate a fost echivalat cu unul fără gheață, la începutul și sfârșitul căruia caracteristicile cantitative ale fitoplanctonului au fost luate egale cu zero. Datele dispariției și apariției gheții în rezervor au fost stabilite conform Serviciului Hidrometeorologic.
Datorită faptului că majoritatea indicatorilor studiați nu sunt măsurați direct, ci sunt calculați pe baza altor măsurători folosind formule adecvate, așa cum nu sunt găsiți
Prin calcularea directă a valorilor medii din rezultatele obținute, este dificil să se determine pur și simplu erorile de reprezentativitate ale acestora folosind metode clasice. Prin urmare, am folosit metoda de evaluare a erorilor indirecte (Zajdel, 1974), în care caracteristica studiată este considerată în funcție de mai multe variabile, a căror acuratețe; ry este predeterminat, iar eroarea sa totală este găsită prin „adunarea pătratică” a erorilor de măsurare ale componentelor individuale folosind calculul diferențial și teoria probabilității. Pe baza acestei metode, au fost compilați algoritmi și programe de calculator originali care fac posibilă evaluarea rapidă a erorilor de măsurare și făcând media rezultatelor obţinute .
III. Caracteristici ale regimului de lumină al studiului
aodosg.sha
Datorită extinderii mari de la nord la sud a teritoriului în care au fost studiate rezervoarele, intensitatea energiei totale a radiației solare care ajunge la suprafața acestora este diferită. Conform datelor medii pe termen lung, valorile sale variază de la 3500 MJ/m1 an în regiunea Kostroma până la 5000 - lângă Astrakhan este de 600, respectiv 750 MJ/m1 lună, la apogeul verii în iunie (URSS Climate Reference Book, 1966).Împreună cu Totuși, în anumite zile ale perioadei de vară (iulie-august) intensitatea energiei solare peste rezervoarele nordice este aceeași !, până la „4 MDch-”./m2 „” ug pp
Ponderea PAR în fluxul de radiații al spectrului general peste corpurile de apă din bazinul Volga Superioară în zilele senine, așa cum se obișnuiește în lucrările hidrobiologice (VbiJenweuier și colab., 1974), este de 46%, dar pe măsură ce tulbureala crește la 57% (Pyrpna, ¡935).
Observațiile privind distribuția radiației solare totale la adâncime în rezervoarele bazinului Volga au arătat că transparența maximă pentru acestea (până la 2 m de-a lungul discului Secchi) este de 1% din totalul.
Cantitatea de radiație eliberată în apă nu este înregistrată la o adâncime mai mare de 2 m. Numai în rezervorul Volgograd, cu o transparență a apei de până la 2,4 m, astfel de valori ale radiațiilor au fost înregistrate la adâncimi de 3-4 m (Rutkovskaya, 1965). În apele lacurilor mai transparente, aproximativ 1% din radiația care intră în apă se observă și mai adânc: la 5-6 m în Lacul Ladoga (Mokievsky, 1968); 6-7 m în Onega (Mokievsky, 1969; Pyrina, 1975(a)); 6-8 m în lac. Pleshcheyevo (Pyrina, 1989(a)). S-a observat o creștere a adâncimii de penetrare a radiațiilor, deoarece înălțimea Soarelui deasupra orizontului a crescut în timpul zilei, precum și spre sud (Rutkovskaya, 1965).
La intrarea în apă, cele mai exterioare razele lungi și scurte se atenuează mai întâi și numai radiațiile apropiate de PAR pătrund mai adânc de 1 metru (Rutkovskaya, 1965; Pyrina, 1965; Pokatilova, 1993). Rolul principal în atenuarea radiației solare în rezervoare îl au detritusurile și suspensiile minerale. Pe acest fond, absorbția luminii de către fitoplancton - așa-numitul „efect de auto-umbrire” (Talling, 1960) - este slab resimțită. Poate fi observat numai vara, când apa „înflorește” cu albastru-verde, care coincide cu o perioadă de stabilitate crescută a masei de apă, când ponderea algelor în cantitatea totală de particule în suspensie devine predominantă (Pyrina, Rutkovskaya, Ilyinsky, 1972). Și numai în apele limpezi ale lacului, cum ar fi lacurile. Pleshcheyevo, influența optică a fitoplanctonului poate fi urmărită destul de clar chiar și în perioada de homotermie (Pyrina, Sigareva, Balonov, 1989).
În apele tulburi ale rezervoarelor se observă valori crescute ale radiațiilor difuze care se întorc din adâncime. Conform măsurătorilor PAR din rezervorul Rybinsk, cu o transparență pe discul Secn de 0,7-1,5 m, ochiul a reprezentat 2-10% din radiația care intră în apă, în timp ce în Lacul Onega - transparență de aproximativ 4 m - mai puțin decât 1% (Pyrina, 1975(a)).
Măsurătorile spectrale ale PAR penetrante în rezervorul Rybinsk și lacul Onega (Fig. 1) au arătat că în stratul de suprafață al apei există o proporție destul de mare de raze roșii-portocalii cu lungimi de undă de peste 600 nm, precum și cele albastre ( X - 380-480 nm). Cu toate acestea, ambele se estompează rapid, iar razele verzi pătrund cel mai adânc (X = 480600 nm). În apele cu o valoare a culorii de peste 70 de grade pe scara platină-cobalt, sa observat o scădere crescută a proporției de radiație albastră (X = 380480 nm) absorbită de stratul superior de apă (Pyrina, 1975(a)) .
Observații de iarnă. asupra pătrunderii PAR prin stratul de zăpadă și zăpadă, efectuată în rezervorul Rybinsk, a arătat că, în condiții tipice, cu un strat de zăpadă de 20 cm și grosimea gheții de 80 cm, nu mai mult de 0,04% din energia radiației primite ajunge. apa (care s-a ridicat la 200-250 W/m"), aceasta se datorează reflexiei intense din zăpadă (80%), proporția de retroîmprăștiere a zăpezii (3%) și radiația întârziată de aceasta (13%) este relativ mic După topirea zăpezii, cantitatea de radiație care pătrunde sub gheață crește la 18-20%,
Orez. I. Penetrarea razelor solare de diferite lungimi de undă în coloana de apă a lacului Onega (a-c) și a rezervorului Rybinsk (d-f), % o g
energia radiațiilor primite.
1 - L = 380-800 im; 2-X-480-800 nm; 3 - L = 600-800 k\<; 4- Л = 680-80") им; вертикаль вниз - прозрачность по белому диску; цифры гмд пен ■ цветность по пяатиново-кобалътовой шкале.
și poate fi urmărit la o adâncime de 2 mega - 0,4%. În același timp, se remarcă reproducerea fitoplanctonului (Pyrina, 1984.1985(a)).
Ținând cont de complexitatea măsurătorilor PAR subacvatice, care necesită echipamente speciale, precum și de faptul că, după trecerea printr-un strat relativ mic de apă, rămâne doar această parte a spectrului solar, am studiat modelele de atenuare a fluxului integral de radiație. într-un rezervor pentru a utiliza datele actinometrice standard ale Serviciului Hidrometeorologic.
Când se descrie în profunzime cursul radiației solare, se folosește pe scară largă legea clasică Bouguer, care, după cum se știe, nu este în întregime legitimă, întrucât legea este valabilă numai în cazul radiațiilor monocromatice. Valorile radiațiilor calculate folosind formula lui Bouguer:
(unde 1r si 10 sunt intensitatea radiatiei la adancimea r si intrate* in apa, si „indicatorul atenuarii verticale), se abat de la cele masurate - in straturile superioare ale apei sunt supraevaluate, in straturile profunde sunt supraestimate. sunt subestimate, ceea ce a fost remarcat într-o serie de lucrări de SDovgy, 1977; Szumiec, 1975; Kirk, 1983). Pentru a compensa această abatere, s-a propus introducerea în exponentul formulei (1) a parametrului r la o putere. mai puțin de 1, în special, egal cu 0,5 (Rosenberg, 1967), iar datele naturale (Are, Tolstyakov, 1969; Larin, 1973) au confirmat posibilitatea de a calcula radiația penetrantă cu această modificare:
Cu toate acestea, aplicarea formulei modificate (2) în studiile de față a relevat o supraestimare semnificativă a intensității radiațiilor calculate pentru orizonturile inferioare, care trebuie luată în considerare la determinarea adâncimii fotosintezei și a valorii integrale a acesteia într-o coloană de apă sub 1. m1. Studiul acestui fenomen a arătat că cea mai adecvată descriere a cursului radiației solare în profunzime se obține prin utilizarea formulei (2) cu o valoare variabilă a exponentului (u) pentru parametrul r,
Valoarea lui n variază în funcție de culoarea apei, iar la valori mici este aproape teoretică. De exemplu, pentru lac. Pleshcheyevo cu culoarea apei pe scara platină-cobalt de 10 grade "=0,6 și pentru
Rezervor Rybinsk cu o culoare de 50 de grade l = 0,8 (Pyrina, 1989).
Astfel, studiul condițiilor de lumină pentru fotosinteza subacvatică din rezervoarele studiate a arătat o gamă largă de adâncimi de penetrare a radiației solare, variind, cu insolație intensă, de la 2 - 5 m în apele slab transparente ale lacurilor de acumulare Volga până la 7 - 12 m. m în lacurile Onega și Pleshcheyevo. Datorită atenuării rapide a razelor extreme de unde scurte și lungi la o adâncime relativ mică, situate în partea superioară! Strat de .5 metri, radiația penetrantă devine identică în spectrul PAR. Datorită atenuării selective de către apă, se observă o abatere a progresiei acesteia în adâncime de la respectarea legii exponențiale, care poate fi compensată matematic. „Acest lucru face posibil, cu suficientă acuratețe în lucrările hidrobiologice, să se calculeze energia iradierii solare necesară pentru fotosinteză din intensitatea radiației solare peste un rezervor, inclusiv cea înregistrată pe uscat de cea mai apropiată stație actinometrică.
Determinările pigmenților de fitoplancton, începute în 1958 în rezervoarele Ivankovo, Rybinsk și Kuibyshev și apoi efectuate în toate corpurile de apă studiate, au arătat o gamă largă de fluctuații ale concentrațiilor acestora (Tabelul 1). Pentru pigmentul principal - clorofila "a", acestea variază de la sutimi de microgram pe litru până la!00 sau mai mult "calculat pentru zona eufotică a rezervorului. Cu toate acestea, fiecare shnoloemo" este caracterizat de un anumit nivel de valori medii pentru sezonul betainei și valorile maxime atinse, în funcție de care pot fi utilizate pentru a judeca asocierea trofică a apelor sale.Conform acestui principiu, rezervoarele aproape tuturor corpurilor de apă au fost acoperite de moștenire - de la lacul monotrofic Onega până la Ivankoch eugrofic Rezervor.
Modificările concentrațiilor de clorofilă în interiorul unui rezervor sunt asociate în principal cu dinamica sezonieră a fitoplanctonului, în perioada de creștere în masă a cărei (de regulă, în mai și iunie ■ !iguete) se observă valorile maxime ale acestora. ] 1a nrig^p^ al lacului de acumulare Rybinsk se arată că înălțimea și momentul acestor maxime în anumite momente depind de caracteristicile hidrometeorologice ale anului (Pyrina, Sigarepa, 1986). Aici am urmărit, de asemenea, uro-ttagt, tsoggceggtrpcpy de clorofilă în sub-gheață gter.tod, când nu a fost livrat mai mult de 0,2 µg/l. Abia după dispariția zăpezii și pătrunderea a aproximativ I MJ/m2 ■ zi de radiație fotosintetic activă sub gheață, concentrațiile de clorofilă au crescut la 0,4 µg/l și a început vegetația fitoplanctonului (Pyrina, 1985(a)).
Tabelul I
Rezervoare ani, clorofilă, sursă
luni informații µg/l
Ivankovskoe 1958, V-IX 12.5 Pyrina, 1966
vdhr. 0 - 2 m 1970, V-X 13,3 Elizarova, 1976
1973-1974, V-X. 26.7-31.8 Pyrina, Sigareva,
1978, V-X 14.2 Pyrina, Sigareva,
■ inedite
Rybinskoe 1958, V-X 6.6 Pyrina, 1966
Raza principală. 1969-1971, V-X 3,4 - 6,7 Elizarova, 1973,
1972-1976, V-X 6.2- 10.0 Pyrina, Sigareva,
1977-1979, V-X 6.6 - Yu.o Mineeva, Pyrina,
1980-1982, V-X 9.3-18.2 Pyrina, Mineeva,
1983-1985, V-X 15,4-19,2 Pyrina, 1991
1986-1990, V-X 9,4-13,8 Ibid.
1991-1993, V-X 12.6-.14.6 Pyrina, inedită.
Kuibyshevskoe 1958, VI-X 7.9 Pyrina, 1966
vdhr. 0 - 3 m
Lacul Alb. 1976-1977, V-X 3,8 -5,0 Pyrina, Mineeva și
0-2 m dr., 1981
Oz. Pleshcheyevo 1983-1985, V-X 6.2-10.0 Pyrina, Sigareva,
Balonov, 1989
Oz. B. Kharbey 1969, VII-VIH 2.0 - Elizarova, Pyri-
pe, Getzen, 1976
Onega 1967-1968, 0,57 - 0,95 Pyrina, Elizarova,
lac. 0 - 5 m VII-VIIÍ Nikolaev, 1973
Lacul Ladoga - 1973-1974, VIII 4,60 Pyrina, Trifo-
ro. 0 - 4, 0 - 5 m nova, 1979
R. Yenisei. 1984-1985, 4,2-7,2 Pyrina, Priyma-
0 m - jos VWX Chenko, 1993
Notă: sunt date valori medii pentru rezervorul și sezonul de vegetație, cu excepția Lacurilor Onega și Lacului Ladoga, unde datele sunt mediate pentru sezonul de vară.
Diferențele în concentrațiile de clorofilă între corpurile de apă sunt mai puțin pronunțate. Doar zonele care primesc ape râurilor îmbogățite cu substanțe nutritive, precum și ape de mică adâncime, în special izolate și slab acoperite cu macrofite, unde conținutul de degradare este crescut (Egshzarova, 1976, 1978; Elizarova, Sigareva, 1976; Elizarova, Pyrina, Getsen, 1976). ; Pyrina, 1978; Pyrina, Pltarova, Nikolaev, 1973; Pyrina, Priymachenko, 1993). Diferențele la scară mică în cadrul unei mase de apă omogene sunt relativ mici și sunt asociate în principal cu acțiunea factorilor dinamici - fluxurile de circulație care determină transportul integral al fitoplanctonului, amestecarea vântului și stratificarea apelor (Pyrina, Sigareva, Balonov, 1989; Pyrina, Mineeva, Sigareva şi colab., 1993).
În majoritatea rezervoarelor studiate, produsele de transformare a clorofilei - fenomene - au fost observate în cantități semnificative. De exemplu, în lacul de acumulare Rybinsk, cantitatea de feopigmenți cociasiiU este de 20-30% din cantitatea cu clorofilă pură, iar la începutul verii minime de fitoplancton se ridică la 60-70%, în special în straturile inferioare de apă (Pyrina). , Sigareva, 1986; Pyrina, Mineeva, 1990). În Lacul Pleshcheyevo în acest moment, aceeași cantitate de feopigmente este observată în stratul de suprafață de 2 metri (Pyrina, Sigareva, Balonov, 1989; Pyrina, 1992), care este combinată cu o abundență crescută de zooplancton ierbivor (Stolbunova, 1989) . Numai în perioada de încălzire maximă a verii, care este de obicei asociată cu „înflorirea” apei albastre-verde, conținutul de feopigmenți este mai mic de 10%. Aceasta coincide cu aportul intens de radiație solară, care împiedică acumularea de clorofilă cu conținut de clorofilă în zona iluminată din cauza efectului distructiv al soarelui (Yentsch, 1965; Moreth Yenisei), 1970), precum și păstrarea înaltului. stabilitatea stratului superior de apă, unde are loc acest proces (Pyrina, Sigareva. 19X6 ).
Alți pigmenți verzi – clorofilele „b” și „c” – au fost găsiți în cantități mult mai mici. Conținutul de clorofilă c" în diatomeele de sud, criptofitele și dinofitele este puțin mai mare (până la 30%) din clorofila totală decât clorofila "b" (câteva procente). Cu toate acestea, apele de coastă, estuare și alte ape sunt îmbogățite cu substanțe nutritive și, în consecință, algele verzi și euglenice - purtători de clorofilă „b”, conținutul acesteia poate ajunge la 10%.
Carotenoizii, măsurați în unități arbitrare, corespund de obicei clorofilei în termeni cantitativi. S-a observat o tendință de creștere a raportului lor cu clorofila, atât în conținut absolut, cât și în raport extinctiv, pe măsură ce productivitatea apei scade. De exemplu, în partea centrală a lacului de acumulare Rybinsk este mai mare decât în cursurile râului și zonele de coastă, iar în partea centrală a acestui rezervor este mai mare decât în rezervorul Ivankovo (Elizarova, 1973, ¡976; Elizarova, Sigareva.1976;Pyrina, Sigareva
1978, 1986). Este chiar mai mare în Lacul Onega, în special în zona sa adâncă, și în tundra lacurile Kharbey (Pyrina, Elizarova, Nikolaev, 1973; Elizarova, Pyrina, Getsen, 1976). Excepție este râul Yenisei, care, cu o productivitate relativ scăzută a apei, se caracterizează printr-un nivel mai scăzut al acestui raport (Pyrina, Priymachenko, 1993). La analizarea raportului pigmenților verzi și galbeni sub aspect sezonier, se observă o creștere la începutul verii minime a fitoplanctonului, în special în timpul reproducerii fitofagelor (Pyrina, Sigareva, Balonov, 1989) și o creștere foarte puternică iarna (Pyrina). , 1985/a).
În ceea ce privește conținutul specific de clorofilă „a” (de obicei 2-5 μg/mg) și carotenoide (1-5 μBri/mg) în biomasa fitoplanctonului, nu sunt detectate diferențe semnificative în corpurile de apă studiate (Pyrina, 1963; Elizarova, 1974, 1976; Pyrina, Elizarova, 1975; Elizarova, Pyrina, Getsen, 1976; Pyrina, Priymachenko, 1993). Doar o creștere a conținutului de clorofilă s-a remarcat în biomasa fitoplanctonului cu o proporție notabilă de alge verzi, ca și în culturile acestor alge (Pyrina, Elizarova, 1971), precum și în comunitatea oligodominantă cu o predominanță de alge mici. specii celulare din genul Biriapsis (Pyrina, Sigareva, Balonov, 1989) ,
Astfel, în timp ce modelele generale ale dinamicii sezoniere și spațiale ale pigmenților fitoplanctonului sunt similare, nivelul concentrațiilor acestora în corpurile de apă studiate variază foarte mult, variind în medie de la 1-2 μg/l în timpul sezonului de vegetație în lacurile neproductive Onega și Bolshaya Kharbey la 30 μg/l - în rezervoarele eutrofice Ivankovsky? Cu toate acestea, conținutul specific de clorofilă în biomasa fitoplanctonului, care este similar în setul de pigmenți și compoziția speciilor dominante, este destul de apropiat. Nu există urme ale unor particularități ale caracteristicilor pigmentului asociate cu adaptarea la lumină a fitoplanctonului din rezervoare optic diferite.Rolul condițiilor de lumină se manifestă numai în reglarea procesului de formare și acumulare a derivaților de clorofilă în zona eufotică a unui rezervor în timpul perioadei. de vreme anticiclonică stabilă.
V, Activitatea fotosintetică a fitoplanctonului
Pe baza valorii producției primare de fitoplancton într-o coloană de apă sub 1 m2, majoritatea corpurilor de apă studiate se încadrează în categoria mezotrofe, deși nivelul de trofeu al acestui tip de apă variază: de la un nivel apropiat de oligotrof pentru tundră. lacul B. Kharbey până la învecinat cu eutrofic pentru rezervorul Rybinsk (Tabelul 2). Doar lacul Onega este clasificat drept rezervoare tipic oligotrofe, iar lacul Ivankovskoe și secțiunea Cheboksary sunt clasificate ca fiind tipic eutrofice. Lacul ocupă un loc aparte. Pleshcheyevo, pentru care
masa 2
Valori medii ale producției primare de fitoplancton în corpurile de apă studiate în perioada de vegetație (g C/m2 zi)
Ivankovskoe 1958 0,90 Pyrina, 1966
vdhr. 1970-1973 0,90 - 1,28 Pyrina. 1978
1974-1976 0,36 - 0,68 Sappo, 1981
1979-1980 0,91 - 1,35 Tarasenko, 1983.
Rezervorul Rybinsk 1955 0,28 Sorokin, 1958
1958 0,54 Pyrina, 1966
1959 0,59 Romanenko, 1466
1001-1905 0,10-0,19 Romanenko, 1985
1966-1971 0,21-0,48 La fel
1972-1973 0,69 - 0,72 La fel
1974-1980 0,18-0,35 La fel
1981 0,83 La fel
1982 0,52 Mineeva, 1990
Gorkovskoe 1956 0,41 Sorokin, Rozanova,
vdhr. Sokolova, 1959
1967 0,76 Tarasova, 1973
1972 0,89 Tarasova. 1977
1974-1979 0,31 - 0,85 Shmelev, Subbopsha. 1983
R. Volga pentru parte - 1966 2,25 Tarasova, 1970
tke Ceboksary-
Kuibyshevskoe 1957 0,66 Salmanov, Sorokin,
1958 0,83 Pyrina, 1966
1965-1966 0,37 - 0,72 Ivatin, 1968, 1970
1967-1971 0,32 - 0,56 Ivatin, 1974, 1983
Saratov 1971-1973 0,32-0,50 Dzyuban, 1975, 1976,
Volgograd 1965-1968 0,30 - 0,63 Dalechina, 1971, 1976
vdhr. 1969-1974 0,33 -0,64 Dalechina, Gerasimo -
1975 1,50 La fel
Continuarea tabelului 2
Rezervoare Ani Sursa primară
cercetarea informațiilor de producție
Lacul Alb 1976-1977 0,21 -0,31 Pyrina, Mineeva și
Oz. Pleshcheyevo 1983-1984 1,36- 1,86 Pyrina, Sigareva,
Dzyuban,)989
Lacul Onega 1966, VII 0,05 - 0,20 Sorokin, Fedorov,
Lacul Ladoga 1973-1974, 0,32-0,60 Pyrina, Trifonova,
Oz. B, Kharbey. 1968-1969 0,14-0,26 Pyrina, Getsen,
Vainshteii, 1976,
R. Ienisei 1984 0,86 Prinmacenko, 1993
Notă: la conversia în carbon, datele metodei oxigenului au fost înmulțite cu 0,375; pentru Lacurile Onega și Ladoga, valorile maxime sunt date pentru partea deschisă
Cele mai mari valori ale producției primare s-au obținut (vara până la! 3 g Og/m2 zi), în ciuda faptului că aici conținutul de clorofilă și intensitatea fotosintezei pe unitatea de volum de apă sunt relativ scăzute.
Cursul sezonier al fotosintezei planctonului în corpurile de apă studiate este caracterizat de modele generale și este determinat în principal de modificări ale abundenței algelor. În lacuri și rezervoare asemănătoare lacurilor, cum ar fi Rybinsk și Kuibyshev, unde periodicitatea modificărilor grupurilor dominante de alge este clar vizibilă, se observă vârfuri de primăvară și vară, deși primul dintre ele este de obicei mai puțin pronunțat din cauza influenței limitatoare. de temperaturi scăzute (Pyrina, 1966; Pyrina, Mineeva și colab., 1981; Pyrina, Sigareva, Dzyuban, 1989), În rezervoare de tip râu cu debit crescut, de exemplu, Ivankovsky, unde diatomeele predomină pe tot parcursul sezonului de vegetație, cursul sezonier al fotosintezei este mai netezit. Cu toate acestea, chiar și aici la mijlocul verii se observă valorile sale maxime (Pyrina, 1978). În apele puțin adânci, în timp ce natura generală a dinamicii sezoniere este similară cu zona de adâncime, a fost observată o schimbare a vârfurilor fotosintezei către date anterioare (Pyrina, Bashkatova, Sigareva, 1976). În timpul iernii, judecând după observațiile individuale în rezervorul Rybinsk (Romanenko, 1971), fotosinteza este extrem de slabă din cauza lipsei de lumină sub stratul de zăpadă și gheață (Pyrina, 1982, 1985(a)).
Diferențele spațiale în producția primară sunt strâns legate de gradul de dezvoltare a fitoplanctonului (conținutul de clorofilă). Cu toate acestea, acest lucru este clar vizibil numai în ceea ce privește intensitatea fotosintezei pe unitatea de volum de apă. Pentru valorile sale integrale, calculate pe unitatea de suprafață a rezervorului, acest model este încălcat, ceea ce este într-un fel sau altul legat de condițiile regimului de lumină.Astfel, în apele puțin adânci bogate în fitoplancton cu adâncimi mai mici decât zona eufotică. , producția primară de iod 1m: este relativ mică datorită stratului fotosensibilizant de limitare din partea inferioară (Pyrina, Bashkatova, Sigareva, 1976; Pyrina, 1978; Pyrina, Sigareva, Dzyuban, IW).În puternic „înflorire” albastru-verde ape, „efectul de auto-umbrire” limitează accesul energiei luminoase la fitoplanctonul orizontului inferior și reduce tinerețea fotosintetică la animalul de companie. Astfel de ape se caracterizează printr-un profil vertical particular al fotosintezei cu un pronunțat, presat la suprafață, stratul maxim.Dimpotrivă, cu pătrunderea adâncă a razelor solare, cum ar fi lacul transparent Pleshcheyevo, precum și unele zone din Volga de Jos, care primesc, în plus și o doză crescută de energie solară, producție primară într-o coloană de apă sub 1 m3 atinge un nivel ridicat cu o biomasă relativ mică de alge (Pyrina, Rutkovskaya, 1976; Gerasimova, 1981; Pyrina, Sigareva, Dzyuban, 1989). In lac În Pleshcheyevo, randamentul crescut al producției primare pe unitatea de suprafață se datorează, de asemenea, caracteristicilor ecologice ale formelor de masă de fitoplancton de vară - alge dinoft din genul Ceraiium, care se pot amesteca activ în căutarea condițiilor optime pentru acesta (Heaney, Talling, 1980). ). Sub radiații solare intense, se acumulează la o anumită adâncime, în zona de fotosinteză maximă, contribuind la creșterea valorilor sale la suta de litri de iod de apă la 1 m; (Pyrina, Sigareva, Dzyuban, 14S9).
Modelele de distribuție a intensității fotosynchronice de-a lungul profilului vertical sunt în general similare pentru toate studiile germane și sunt determinate în principal de cantitatea de pergestie la o anumită adâncime. Foyueshpe maximă! observată la adâncimi la care iradierea este de aproximativ 4,0 MD-k/m2 zi PAR, iar atenuarea sa este la 0,02 MJ/u3 ■ zi. În zilele senine, astfel de condiții sunt create la adâncimi, respectiv, 1-2 și 10 m până la t.i Pleshcheyevo ("Pyrina, Sigareva. Dzyuban, 1989). 1,0-1 5 n 1-9 m i, Lacul Ong-ch (Sorokin . Fedor ")", iУ69>, Ö-0,5 și 3-6 m și lacul L. Kh.<(" vi* (üupiuu, ;сцсн, Ьакншчсин, 1976), 0-0,4 и 2-5 м в волжских водохранилищах (Пырина, Рутковская, 1976). Так, практически одна и та же энергия облученности (96 кал или 4,02 МДж на I м2 в сутки) наблюдалась на глубинах максимального фотосинтеза в Рыбинском вопохпл-ннлнще как весной, при интенсивной вегетации диатомовых, так и ж-(ом, когда в массе развивались сине зеленые (рис. 2). Однако в одном и том же водоеме эти, соответствующие характерным параметрам фото-синтезнрующей зоны, глубины могут значительно варьировать в шви-
Orez. 2. Distribuția pe verticală a radiației solare (X = 380-800 nm) și fotosinteza fitoplanctonului cu predominanță de diatomee sau alge verzi de zăpadă. Rezervorul Rybinsk, 1972
a - „înflorirea” diatomeelor (16 mai), b – „înflorirea” sinergiei (7 iulie). 1 - fotosinteza in mg O/l.zi, 2 - radiatii e MJ/m "supa redusa la scara MRE (Scrisoare metodologica... ¡982, citata din: Evnevich, 1984). Sagetile indica energia radiatiei la adâncimea fotosintezei maxime .
simulare a influxului de radiație solară, ceea ce duce la o modificare a valorilor integrale ale producției primare sub 1 mg (Pyrina, Mineeva, în presă).
Eficiența utilizării energiei solare în timpul fotosintezei fitoplanctonului în rezervoarele studiate variază de la 0,01-i% din spectrul integrat de radiații incidente pe apă, sau 0,022% din energia PAR. Cele mai mari valori (aproximativ 1%) au fost obținute în zona lacului de acumulare Cheboksary, în rezervoarele Kuibyshev și Ivankovsky, caracterizate prin fitoplancton abundent (Pyrina, 1967 (a), 1978), precum și în lac. Pleshcheev - datorită adâncimii mari a zonei fotosintetice (Pyrina, Sigareva, Dzyuban, 1989). Aceste valori corespund celor mai mari cunoscute pentru ape dulci foarte productive (Talling et al., 1973). Folosind exemplul Lake Pleshcheyevo a urmărit influența reciprocă a indicatorilor regimului de lumină și a cantității de fitoplancton în procesul de utilizare a luminii solare, care crește proporțional cu concentrația de clorofilă sau 10 μg/l și scade pe măsură ce cresc în continuare și manifestarea „ efect de auto-umbrire” (Pyrina, Sigareva, Dzyuban, 1989).
Relația dintre intensitatea fotosintezei la maxim de-a lungul profilului vertical și biomasa algelor sau clorofilei conținute în acestea, care caracterizează activitatea de asimilare a fitoplanctonului, variază în limite destul de apropiate în toate corpurile de apă. În cele mai multe cazuri, raportul dintre fotosinteză și biomasă (coeficientul de activitate a biomasei) este de 0,3-0,4 mg Qi/mg, iar clorofilă (număr de asimilare) - 0,10-0,20 mg Og/µg pe zi. Valorile lor crescute sunt tipice pentru estuar, ape puțin adânci și alte zone, în fitoplanctonul cărora algele verzi reprezintă o proporție semnificativă (Pyrina, 1959 (a), ¡967 (5), 1978; Pyrina. Trifonova. " " 79; Pyrina şi colab. 1981); Sigareva, 1984; Pyrina, Sngya-ro"i, Baloiov, 1989).
Studiile țintite ale producției primare arată că valorile sale maxime de-a lungul profilului vertical sunt determinate de cantitatea de plancton foying și de clorofila pe care o conține, iar valorile integrale pentru coloană sunt determinate de caracteristicile regimului de lumină, care determină adâncimea de răspândire a fotosintezei în rezervor.Ca urmare, în ape transparente slab colorate, absorbind puțină energie radiantă, a trimis ■ a mâncat randamentul fotosintezei pe unitatea de clorofilă și radiația solară care intră.Factorul de lumină determină diferențele semnificative în valorile medii anuale ale producției primare asociate cu durata perioadei de aport intens de energie solară și adâncimea pătrunderii acesteia, în funcție de înălțimea Soarelui deasupra orizontului.
VI. Oceica dependențelor de lumină ale fotosintezei fitoplaic-
În experimente pentru a studia activitatea fotosintetică a fitoplanctonului în funcție de energia radiației solare subacvatice, efectuate într-o zonă mare de apă din Volga de mijloc și inferioară în rezervorul Rybinsk și în lac. Pleshcheev, a fost obținută o serie de curbe luminoase tipice ale fotosintezei. După normalizare” conform valorii sale maxime pentru fiecare probă, luată ca 100%, datele din diferite experimente, stabilite în condiții similare de iradiere, s-au dovedit a fi grupate destul de strâns, atât în ceea ce privește energia PAR cât și spectrul general solar ( Fig. 3).doar unele dintre discrepanțele lor depind de compoziția grupelor dominante de alge: pentru fitoplanctonul cu predominanța verde, punctele au fost deplasate spre energie de iradiere mare, iar pentru fitoplanctonul din diatomee și, în special, albastru-verde. alge - în regiunea valorilor sale inferioare (Pyrina, 1967; Pyrina, Rutkovskaya, 1976).
Astfel de rezultate sunt în concordanță cu modelele de distribuție verticală a algelor bentonice în rafturile mării, unde reprezentanții verzi cresc mai des în condiții de lumină puternică la adâncimi mici, în timp ce diatomeele, maro și, în special, cele roșii pătrund mai adânc. Un fenomen similar a fost observat în unele corpuri de apă dulce de adâncime (Gessner, 1955; Kirk, 1983). Semne de adaptare la lumină asociate cu capacitatea de absorbție a luminii a pigmenților au fost observate și în fitoplanctonul marin crescut de la diferite adâncimi (Koblenz-Mishke, 1980). Cu toate acestea, în experimentele cu culturi (Curtis, Juday, 1937; Ichimura, Aruga, 1958; și altele), inclusiv izolarea planctonului din rezervorul Rybinsk (Pyrina, 1959), nu au existat modele similare în răspunsul algelor la SVST. în funcţie de setul de pigmenţi s-a trasat.
Ținând cont de această inconsecvență, precum și de faptul că fitoplanctonul din Volga, ca și alte rezervoare studiate, este reprezentat în principal de diatomee și verzi de zăpadă, care s-au dovedit a fi destul de apropiate în relația lor cu lumina, o singură curbă de lumină a fost construit pentru totalitatea rezultatelor tuturor experimentelor efectuate în rezervoarele Volga (Fig. 3). În acest caz, am folosit date despre energia spectrului general, care sunt cele mai accesibile pentru o gamă largă de cercetători. Aproximativ, adică Fără a lua în considerare fotoinhibația, care este relativ puțin exprimată în rezervoarele Volga, dependența fotosintezei fitoplanctonului de radiația solară care pătrunde în apă a fost descrisă printr-o ecuație de forma:
Az"=Iz/(0,35 + 0,009 l.J, (4)
5 10 50 yua)50 ¿50 /Gal!cis
Radiații de iederă pe zi
Orez. 3. Viteza relativă de fotosinteză a fitoplanctonului în rezervoarele Volga la diferite intensități de intrare a radiației solare (spectru general).
1,2 - fitoplancton cu predominanță de alge albastru-verzi sau respectiv diatomee; 3 - fitoplancton mixt cu predominanţă a celor verzi. Fotosinteza în % din maxim de-a lungul profilului vertical; energia radiației solare în MD/s/m cabană-pe scara MRE.
unde 1g și Az"" energia radiației și intensitatea fotosintezei (în% din maximul de-a lungul profilului vertical) la adâncimea r; Ar -(AJAMaKC) 100, unde Ar și Amax sunt în unități absolute. Ecuația rezultată (4) este similară cu binecunoscuta ecuație Michaelis-Menten, care poate fi considerată cea mai simplă dintre opțiunile comune pentru descrierea efectului luminii asupra fotosintezei. În special, este o variație a ecuației Bannister (Bannister, 1979):
A,=AMaKC!/(Ikm+Im) Ш (5)
(aici A j și Amax sunt intensitatea fotosintezei la energia radiației / și, respectiv, maximă de-a lungul profilului vertical; /^ ■ constantă), care este destul de universală și la t -I se transformă în ecuația Michaelis-Menten, iar la t = 2 în ecuația Smith (Broth, 1994). Acesta din urmă, după cum se știe, formează baza modelului Talling, adoptat pentru calcularea producției primare integrale într-o coloană de apă la 1 mg (Tailing, 1957; Vollenweider, et al., 1974). Variind valoarea parametrului m, este posibil să se descrie curbele de lumină cu o regiune de saturație la o intensitate luminoasă mai mare (de exemplu, dacă m - 2) sau scăzută (»u = 3).
Astfel, ecuația derivată (4) corespunde curbelor de lumină ale fotosintezei cu o pantă destul de plată, conducând la o deplasare a maximului către regiunea de iradiere crescută și o manifestare slabă a fotoinhibiției. Este posibil ca acest tip de dependență să fie legat într-o oarecare măsură de particularitățile iradierii din experimente, care s-au schimbat în termeni spectrali pe măsură ce a scăzut cu adâncimea. Și totuși, lipsa de adaptare pronunțată la lumina slăbită de apă poate fi, evident, considerată caracteristică pentru cea mai mare parte a fitoplanctonului din corpurile de apă studiate, unde factorul de amestecare este semnificativ și celulele de alge nu trebuie să rămână în straturi adânci de apă pentru o perioadă lungă de timp. timp.
VII. Rolul caracteristicilor regimului de lumină al diferiților ani în variabilitatea interanuală a productivității fitoplanctonului
Folosind exemplul rezervorului Rybinsk, pentru care s-a obținut cea mai completă serie de date privind conținutul de clorofilă, au fost luate în considerare modificările nivelului productivității fitoplanctonului într-un plan pe termen lung cu o evaluare diferențiată a rolului factorilor antropici și climatici, în special, regimul radiaţiei solare (Pyrina, 1991).
Analiza acestor date a arătat o creștere a conținutului mediu de clorofilă din rezervor în perioada fără gheață, începând cu anul 197! g. (Fig. 4). În anii 80, a atins un nivel critic (10 μg/l), dincolo de care zoozoarul este de obicei clasificat ca eutrofic. Aceasta a coincis cu o creștere a cantității de nutrienți, în special a fosforului total, ale cărui concentrații în rezervorul principal au crescut de la 33-47 µg/l în 1965 și 1970. (în medie pentru anotimpurile individuale) până la 40-60 µg/l în 1980 (Bylinkina, Trifonova, 1978; Razgulin, Gapeeva, Litvinov, 1982). Ele intră în principal prin tronsonul Sheksninsky, care primește apele uzate din complexul industrial Cherepovets, așa cum indică o creștere deosebit de accentuată a concentrațiilor de clorofilă în această secțiune a rezervorului.
Pe fondul unei tendințe generale de creștere s-au remarcat creșterile mari ale concentrațiilor de clorofilă, corespunzătoare unui focar de vegetație fitoplanctonică în ani cu vreme anticiclonică (1972, 198!, 1984), care s-au datorat afluxului intens de radiație solară: 15. -16 MJ/m* zi. în medie pentru sezonul de vegetație (mai - octombrie) cu cele mai frecvente valori în lunile de vară variind între 20-25 MD/m2 ■ zi. Apoi, conținutul de pigment a scăzut ușor, dar a rămas mai mare decât în anii premergătoare erupției și a rămas în jurul acestui nivel timp de câțiva ani, până când a început următoarea perioadă de creștere a energiei radiației solare care pătrunde pe Pământ (Fig. 4). În lacul și lacul Ivankovskoe din apropiere. În Pleshcheyevo în acești ani (1972, 1973, 1984), s-au observat, de asemenea, valori crescute ale conținutului de clorofilă (Tabelul 2) și producția primară de fitoplancton (Tabelul 3). O creștere a nivelului producției primare a avut loc și în alte rezervoare Volga în ani similare condițiilor hidrometeorologice, de exemplu în 1975 (Tabelul 3). In lac Windermnr a relevat o influență pozitivă a radiației solare asupra formării biomasei de vârf a diatomeelor de primăvară (K"ea! e1 a!., 1991), Adevărat, aceasta (.kiyu a afectat nivelul mediu anual al biomasei fitoplanctonului (conținutul de clorofilă), care în aceeași perioadă de ani în a fost în mare măsură determinată de alți factori (TaShiv, 1993).
O creștere semnificativă a productivității fitoplanctonului în anii cu vreme anticiclonică se explică prin influența unui set complex de condiții necesare pentru fotosinteză și creșterea biomasei de alge. Datorită aportului intens de radiație solară, adâncimea de răspândire a fotosintezei crește și, în consecință, valoarea sa integrală în coloana de apă pe unitatea de suprafață a rezervorului crește. În condițiile de vânt slăbit caracteristice acestor ani, are loc o încălzire crescută a coloanei de apă și stabilitatea acesteia crește. Acest lucru promovează rezidența pe termen lung a algelor în zona iluminată cu un conținut ridicat de clorofilă activă (fără derivați), fotosinteza și creșterea acestora. Este posibilă stimularea ambelor procese cu elemente biogene, praf
17 16 15 N 13 12 I 10 9 V 7 6
1958 1971. 1975 1979 1983 1987 1991
1969 197E 1977 1901 1985 1989 1993 Ghana
Fig.4. Modificări pe termen lung în intensitatea radiațiilor primite și a conținutului de clorofilă în rezervorul Rybinsk
solar
/ - energia radiației solare totale (spectru total), MJ/m1 zi, în medie mai-octombrie conform Observatorului Hidrometeorologic Rybinsk; 2 conținut mediu de clorofilă „a” (împreună cu phgopigmenți) în perioada lipsită de gheață, μg/l, în stratul superior de apă de 2 metri conform observațiilor la stațiile standard ale lacului de acumulare principal.
distruse prin distrugerea bacteriană a materiei organice, care este, de asemenea, intensificată de căldură (Romanenko, 1985). Evident, în perioada de radiație solară intensă, ecosistemul rezervorului este îmbogățit cu energie utilizată în timpul fotosintezei, iar procesele de viață ale planctonului vegetal sunt de fapt stimulate de nutrienți; asigurând creșterea biomasei sale.
Rezultatele obținute relevă principala condiție pentru funcționarea sistemelor de apă - afluxul obligatoriu de energie solară pentru menținerea acesteia. Din acest punct de vedere, lumina joacă un rol nu mai puțin important decât elementele biogene în reglarea productivității fitocenozelor planctonice și a rezervorului în ansamblu, inclusiv în procesul de eutrofizare a acestuia. Astfel, soluția la această problemă globală este asociată cu evaluarea efectului fstor-ului luminii.
Concluzie
Principalele rezultate ale cercetării sunt următoarele:
1. În funcție de transparența apei, adâncimea de penetrare a razelor solare în rezervoarele studiate variază foarte mult, variind de la 2-5 m în rezervoarele Volga până la 7-12 m în lacurile Onega și Pleshcheyevo sub insolație intensă. Datorită atenuării razelor extreme de unde scurte și lungi „în chiar stratul de cola la o adâncime mică de aproximativ 0,5-1,5 m, radiația penetrantă devine identică în spectrul PAR. În cadrul PAR extinzându-se mai adânc. , razele” din regiunea albastră a spectrului sunt cel mai intens atenuate, mai ales cu culoarea crescută a fundului, apoi roșu, în timp ce verdele pătrunde până la adâncimea maximă. În eanz” cu atenuarea selectivă a radiației sale, codul său de adâncime se va abate aceasta din legea exponentiala corespunzatoare.Abaterea poate fi compensata matematic, ceea ce face posibila calcularea energiei de iradiere necesara fotosintezei subacvatice din intensitatea radiatiei solare peste un corp de apa, inclusiv cea inregistrata de cea mai apropiata statie actmstrncheskon.
2. Conținutul de pigment principal al plantei - clorofila „a” variază de la 1-2 μg/zi b în medie pentru perioada de tegație în apele neproductive ale lacului Onega și lacului tundra. B. Kharbey până la 30 µg/l în rezervorul eutrofic Ivankovo cu valori maxime în timpul sezonului de creștere în masă a fitoplanctonului de la 1,5-3 µg/l până la 100 sau, respectiv, mai mult. Unicitatea dinamicii sezoniere a conținutului de clorofilă în fiecare grup de corpuri de apă, care diferă în ceea ce privește nivelul conținutului de pigment, poate fi urmărită: un vârf ascuțit și vârfuri de vară relativ slab exprimate în Lac. Pleshchee
în, de dimensiuni egale - în Lacul Alb, primăvară, vară și toamnă (sau vară-toamnă) - în rezervorul Rybinsk, o vară - în rezervorul Ivankovskoye, precum și în lacurile tundra Kharbey și, se pare, lacul Onega . Iarna, cu o absență aproape completă a luminii sub un strat gros de zăpadă și gheață, concentrațiile de clorofilă sunt apropiate de 0. Modelele de variabilitate spațială a conținutului de clorofilă în toate corpurile de apă sunt destul de asemănătoare și sunt asociate în principal cu condițiile de regimul hidrodinamic. Nu s-au constatat diferențe semnificative în conținutul specific de clorofilă, precum și raportul acestuia cu alți pigmenți pentru fitoplanctonul algelor dominante omogene ale diferitelor corpuri de apă, precum și semne de adaptare la lumină a sistemului său pigmentar în ape optic diferite și într-un mediu sezonier. aspect. În același timp, se relevă rolul limitativ al radiației solare în procesul de formare și acumulare a derivaților clorofilei, datorită căruia se menține un conținut ridicat al fondului său activ în perioadele cele mai favorabile fotosintezei cu vreme anticiclonică. În anii cu predominanța unor astfel de perioade, se observă o creștere a nivelului general al conținutului de clorofilă din rezervor.
3. Nivelul valorilor producției primare calculate pe unitatea de volum de apă (la maxim de-a lungul profilului vertical) este determinat de cantitatea de plancton de fotosinteză (conținutul de clorofilă) și integral în adâncime - de caracteristicile regimului de lumină , care determină puterea zonei de fotosinteză a rezervorului. Ca urmare, în apele transparente de culoare scăzută, care absorb o cantitate relativ mică de energie radiantă, randamentul producției primare de plancton pe unitatea de clorofilă și energia solară care intră în rezervor crește. Folosind exemplul rezervoarelor din cascada Volga , o creștere a valorilor medii anuale ale producției primare de fitoplancton spre sud datorită aportului crescut de radiație solară.Dimpotrivă, în lacurile tundra, producția primară este limitată de lipsa energiei luminoase datorită intrării limitate a acesteia în rezervorul la altitudini solare scăzute deasupra orizontului și durata scurtă a perioadei fără gheață.
4. Studiul dependențelor de lumină ale fotosintezei a arătat asemănări în răspunsul comunităților de diatomee și alge albastre-verzi tipice rezervoarelor studiate la intensitatea iradierii subacvatice. S-a găsit o reprezentare analitică a acestei relații, care a făcut posibilă îmbunătățirea metodei de calcul pentru determinarea producției primare.
5. Datele pe termen lung privind conținutul de clorofilă obținute în rezervorul Rybinsk arată că, pe fondul fluctuațiilor interanuale, are loc creșterea acesteia, indicând e-
rezervor de apă trofeu. Acest lucru coincide cu o creștere a încărcăturii de nutrienți din partea dezvoltată industrial a coastei. Cu toate acestea, creșteri puternice ale concentrațiilor de pigment în ani cu predominanță
vremea de tip anticiclonic face să creadă despre accelerație -----
acest proces cu o aprovizionare sporită de energie solară și astfel luați în considerare lumina, alături de elemente biogene^! ca factor de eutrofizare a rezervorului.
Studiile efectuate arată cuprinzător rolul important al luminii în asigurarea activității fotosintetice a fitoplanctonului și formarea producției primare a acestora. De asemenea, este important ca energia radiației solare care pătrunde într-un rezervor să poată fi înregistrată folosind metode fizice precise, precum și modelare matematică, la fel ca procesul de fotosinteză subacvatică în sine. În plus, în raport cu fitocenozele planctonice, caracterizate printr-o structură relativ simplă cu interacțiune strânsă cu habitatul, modelele sunt destul de simple și adecvate mediului natural. Toate acestea extind posibilitățile de măsurători instrumentale și automate în studiile de productivitate a ecosistemelor acvatice.
Literatură
1. Aleknn O.A., Semenov A.D., Skopintsev B.A. Ghid pentru analiza chimică a apelor terestre // Leningrad, 1973.
2. Sunt F.E., Tolstyakoo D.N. Despre pătrunderea radiației solare în apă // Meteorol. hidrol. 1969, N 6. P. 53-64.
3. Bell L.N., Chyora S.N., Korniliev V.P. Dispozitiv pentru determinarea cantității de radiații (fotointegrator) // Physiol. rzst, 1959. T.6, numărul. 4. p. 504-507.
4. Berezkin V.A. Aplicarea filtrelor în actinometrie și metode de prelucrare a observațiilor cu acestea // Geophyeich. Sâmbătă, 1932. T.5, numărul E.
5. Bulion V.V. Modele de producție primară în ecosistemele limnice. Sankt Petersburg, 1994.
6. Bylinkina A.A., Trifonova N.A. Elemente biogene // Volga și viața ei. L., 1978. p. 80-92.
7. Vinberg G.G. Experiență în studiul fotosintezei și respirației în masa de apă a unui lac. Pe problema echilibrului materiei organice. Mesaj 1 //Tr. Limnol. Artă. în Kosin. 1934. Emisiunea. 18. P. 5-24.
8. Vinberg G.G. Conținutul de clorofilă ca indicator al dezvoltării cantitative a fitoplanctonului // Al treilea ecologic. conf. Rezumat al raportului Partea 4. Kiev, 1954.. P. 70-74.
9. Vinberg G.G. Producția primară de rezervoare. Minsk, 1960.
10. Vinberg G.G., Sivko T.N. Determinarea conținutului de clorofilă în plancton II Izv. Academia de Științe a BSSR. 1953, N 3. P. 61-74.
11. Vinberg G.G., Sivko T.N., Kovalevskaya R.Z. Metode de determinare a conținutului de clorofilă în plancton și unele rezultate ale aplicării acestora // Producția primară a mării și apelor interioare. Minsk, 1961. p. 231-240.
12. Gerasimova N.H. Dinamica producției de fitoplancton în rezervoarele Saratov și Volgograd conform datelor pe termen lung II Sat. științific tr. Institutul de Cercetare a Lacului și discursuri. peşte x-va. 1981, N 168. P. 65-S1.
13. GOST 17. 1.04.02.-90. Apă. Metodologia pentru determinarea electrofotometrică a clorofilei „a” //M., 1990.
14. Dalechina I.N. Producția primară și distrugerea materiei organice în lacul de acumulare Volgograd în anii 1965-1967. II Tr. Departamentul Saratov GosNIORH. 1971. T.10. pp. 47-59.
15. Dalechina I.N. Producția primară și distrugerea materiei organice în rezervoarele Volgograd în 1968 // Rezervorul Volgograd. Saratov, 1976. p. 55-58.
16. Dalechina I.N., Gerasimova N.A. Fitoplancton și produsele sale tt Dezvoltarea piscicolă și potențialul de bioproducție al lacului de acumulare Volgograd. Saratov, 1980. p. 31-38.
17. Dzyuban A.N. Producția primară și distrugerea materiei organice. substanţe din apa lacului de acumulare Saratov în 1971. - // Biologie internă. apă, Informați. Buletin 1975. N 25. P. 12-15.
18. Dzyuban A.N. Producția primară și distrugerea materiei organice în apa lacului de acumulare Saratov în anul 1972. II Biologia internă. apă Informa. Buletin 1976. N 31. P. 13-16.
19. Dzyuban A.N. Producția primară, distrugerea materiei organice și a numărului de bacterii din apa lacului de acumulare Saratov // Gidrobiol. revistă 1977. T. 13, N 1. P. 81-85.
20. Dovgy T.N. Radiația solară subacvatică pe lacul Baikal. Novosibirsk, 1977.
21. Evnevici.T.V. La calcularea caracteristicilor transparenței atmosferice // Meteorol. hidrol. 1984. N 5. P. 103-109.
22. Elizarova V.A. Compoziția și conținutul pigmenților vegetali din apele hidrobiolului Rybinsk Reservoir II. revistă 1973. T.9, N2. pp. 23-33.
23. Elizarova V.A. Conținut de pigmenți fotosintetici pe unitatea de biomasă fitoplanctonică din rezervorul Rybinsk // Flora, fauna și microorganismele din Volga. Rybinsk, 1974. p. 46-66.
24. Elizarova V.A. Conținutul pigmenților de fitoplancton din rezervorul Ivankovo, conform observațiilor din 1970 // Biologia, morfologia și sistematica organismelor acvatice. L., 1976. P. 82-90.
25. Elizarova V.A. Dinamica sezonieră și distribuția pigmenților de fitoplancton în rezervorul Rybinsk // Biologia și sistematica organismelor inferioare. L., 1978. S. 103-124.
26. Elizarova V.A., Sigareva L.E. Conținutul de pigmenți de fitoplancton în apele de mică adâncime ale lacului de acumulare Rybinsk II Regimul hidrobiologic al apelor de mică adâncime de coastă din lacurile de acumulare Aolga superioară. Yaroslail, 1976. p. 133-147,
27. Zydol A.M. Erori în măsurătorile mărimilor fizice. L., 1974.
23. Ivatsh" A.B. Caracteristicile microbiologice ale rezervorului Kuibyshev în jurul anului 1965. Și Microbiologie, 1968. T.37, nr. 2. p. 360-366.
29. Ivatin A.B. Producția primară și distrugerea materiei organice în sistemul de sănătate Kuibyshetsk în „8ob g.// biolgich gnutr.” sod. Inform. s>yul. 1970. M 8, pp. 15-19.
30. Ivatin A.B. Producția de fitoplancton și distrugerea materiei organice în rezervorul Kuibyshev // Gidrobiol. revistă 1974. T.10, N 3. P. 65-69.
31. Ivatin A.B. Producerea și distrugerea materiei organice și a rezervorului Kuibyshev. L., 1983. P. 90-102.
32. Kirillova T.V. Regimul de radiație al lacurilor și rezervoarelor. L., 1970.
33. Koblenz-Mishke O.I. Absorbția și eliminarea semințelor
n procesul de fotosinteză a fitoplanctonului Mării Negre N Ecosistemele Mării Negre. M., 1E30. p. 132 -139.
34. Ksblznts-Uishye O.I., Voznik V., Ochakovsky Yu.E. (ed.). /p&osik* solar energy z process* foshsknteza chgomomsrsk%:g.. ■l -¿Altai fpuplaneten. M., IS85.
25. „.Soblenz-Mishkp O.I., Vogiyak B., Konooanoye O.V. Md“-P17sg.;;”adică.. r&sy-." -gchnch^och produse // Oi.vawrewm Baptik"! . > „non-isna 1984. N., .037. pp. 385-40/
l. Larin D.A. Despre calculul morții lui Sopntsp, pătrunzând în üiyöb ßQAOSiAoa.7 üöcihhk MSU. Seria V. Geografie. 1973. N 1. S. 102-106.
37. Msweftß "H.f"1. Perps.chnpya producție de platlon FWHihghov g.ndohrlnchlishcha // suport și croduttnod pzd&gich&mmkh și fitocenoze litorale ale rezervoarelor din bazinul Volga. L., 1990. S. 207-216.
38. Mokieasky K.A. Pătrunderea energiei radiante a Soarelui în masa de iod a Lacului Ladoga // Oyasim termic al Lacului Ladoga. L., 1968. P. 73-99.
39. Mokiovsky K.A. Bilanțul de radiații al lacului Onega II Regimul termic al lacului Onega. L, 1973. P. 52-134.
40. Nichilorovich A.A. Fotosinteza și teoria obținerii de randamente mari. XV lectură Timiryazev. M., 1956.
41. Nichilorovich A.A. Activitatea fotosintetică a plantelor ca bază a productivității lor în biosferă și agricultură // Fotosinteza și procesul de producție. M., 1988. P. 5-28.
42. Pokatilova T.N. Atenuarea spectrală a radiației solare în apele naturale II Hidrologia Baikalului și a altor corpuri de apă. Novosibirsk, 1984. p. 14-19.
43. Pokatilova T.N. Utilizarea unui piranometru pentru a măsura energia radiației solare subacvatice // Probleme metodologice în studierea producției primare de plancton în corpurile de apă interioare. Sankt Petersburg, 1993. p. 139-146.
44. Priymachenko A.D. Producția fotosintetică a fitoplanctonului și starea trofică a râului. Yenisei // Studii productive și hidrobiologice ale Yenisei. Novosibirsk, 1993. p. 149-161.
45. Razgulin S.M., Gapeeva M.V., Litvinov A.S. Echilibrul de nutrienți și ioni în rezervorul Rybinsk în 1980. II Studii hidrochimice ale rezervoarelor Volga. Rybinsk, 1982. p. 61-91.
46. Rosenberg. G.V. Fundamentele fizice ale spectroscopiei substanțelor care difuzează lumina // Progrese în fizică, științe. 1967. T.91, N 4. P. 569608.
47. Romanenko V.I. Caracteristicile proceselor microbiologice de formare și distrugere a materiei organice în rezervorul Rybinsk II Producția și circulația materiei organice în rezervoarele interioare. M.-L., 1966. P. 133153.
48. Romanenko V.I. Producția de materie organică de către fitoplancton în rezervorul Rybinsk // Gidrobiol. revistă 1971. T.7, N4. pp. 5-10.
49. Romanenko V.I. Procese microbiologice de producere și distrugere a materiei organice în corpurile de apă interioară. L., 1985.
50. Rutkovskaia 8.A. Caracteristicile proprietăților de radiație ale lacurilor, rezervoarelor și mărilor I/Cercetarea meteorologică marină. M..19S2. p. 117-132.
51. Rutkovskaya V.A. Penetrarea radiațiilor de sare în corpurile de apă ale uscatului și mării II Studii ale circulației atmosferice și ale stratului de aer de suprafață peste Oceanele Pacific și Indian. M., 1965. S. 245-274.
52. Salmanov M.A., Sorokin Yu.I. Producția primară a lacului de acumulare Kuibyshev // Izv. Academia de Științe a URSS, ser. biol. 1962. N 4. P. 603-613.
53. Sappo L.M. Producția primară de materie organică în rezervorul Ivankovo în anii 1974-1976. // Sat. științific tr. Institutul de Cercetare a Lacului și vorbire peşte x-va. 1981. N 105. p. 30-34.
54. Semenchenko!?.A. (Meleshko I.E., Kirmalov R.L., Vladimirova I.A. Spectrometru de înregistrare subacvatică cu telecomandă // Meteorol. hidrol. 1971. N 4. P. 9499.
55. Sigarvva L.E. Conținutul și activitatea fotosintetică a clorofilei în fitoplancton din Upper Zolga. Insulta. Ph.D. biol. Sci. Borok, 1984.
56. Sorokin Yu.I. Producția primară de materie organică în coloana de apă a rezervorului Rybinsk // Tr. biol. Artă. — Borok. 1958. Vol. 3. pp. 66-68,
57. Sorokin Yu.I., Rozanova E.P., Sokolova G.A. Studiul producției primare în lacul Gorki folosind Sm //Tr. Toate hidrobiol. despre-va. 1959. T.9. p. 351-359.
58. Sorokin Yu.I., Fedorov V.K. Determinarea producției primare și a distrugerii materiei organice în Lacul Onega // Fiziologia organismelor acvatice și rolul lor în ciclul materiei organice. L., 1969. P. 4-8.
59. Manual despre clima URSS. L., 1966. Vyl.8, partea 1; emisiune 12, partea 1; numărul 13, partea 1; Problema 29, 4.1.
60. Stolbunova V.N. Zooplancton // Ecosistemul lacului Pleshcheyevo. L., 1989. p. 156-173.
61. Tarasenko L.V. Caracteristicile producției de fitoplancton în rezervorul Ivankovo în anii 70. M., 1963. (Manuscris depus în VINITI 8 iunie 1983 N 3153-83 Depozit)
62. Tarasova T:N. Producția primară și distrugerea materiei organice din râuri. Volga pe traseul de construcție a centralei hidroelectrice Ceboksary în 1956.” // Uch. zap. Gorkovsk. un-ta. Ser. biol. 1970. Problemă. 105. p. 32-36.
63. Tarasova T.N. Producția primară, bacterioplancton și distrugerea materiei organice în rezervorul Gorki și Gedrobiol. judecător 1973. T.9, N 3. P. 5-11.
64. Tarasova T.N. Producția primară și participarea bacteriilor la distrugerea materiei organice din partea lacului din rezervorul Gorki // Ciclul materiei și energiei în rezervoare. Elemente biotice circulaţie. Zada pe Baikal. 1977. p. 69-73.
65. Chekhin L.P. Regimul de lumină al rezervoarelor. Petrozavodsk. 1987.
66. Shmelev G.A., Subbotina E.V. Producția fotosintetică și distrugerea materiei organice în rezervorul Gorki pentru anii 1974-1979. // Sat. lucrări Gork., Volzhsk și Rybin, hidrometeorol. obs. 1983. N 16. P. 51-59.
67. Yanishevsky Yu.D. Instrumente actinometrice și metode de observare cu acestea. L., 1957.
68. Bannister T.T. Descrierea cantitativă a creșterii algelor în stare de echilibru, saturate cu nutrienți, inclusiv adaptarea It Limnoi. Oceanogr.
1979. Vol. 24, N 1. P. 76-96.
69. Burkholder P.R., Burkholder L.M., Rivera J.A. Clorofila „a” la unele corali și plante marine // Natura. 1959. Vol. 183, N 4671. P. 1338-1339. > "
70. Curtis J.T., Juday C. Photosynthesis of algae in Wisconsin Lakes // intern. Rev. ges. Hidrobioi. 1937. Bd. 35, H.2. S. 122-133.
71. Ganf G.G. Producția fotosintetică și relațiile radiație-fotosinteză ale fitopianctonului dintr-un lac ecuatorial de mică adâncime (Lacul George, Uganda) // Oecologia. 1975. Vol. 18, N 3. P. 165-183.
72. Ganf G.G., Heaney S.I., Cony J. Absorbția luminii și conținutul de pigment în populațiile și culturile naturale ale unei cianobacterie non-gaz vacuolate Oscillatoria bourretlyl (Tychomema bourrelyl) și J. Plankton Res. 1991. Vol. 13, N5. P. 1101-1121.
73. Gessner F. Hydrobotanik. Bd.1. Berlin, 1955.
74. Heaney S.4., Tailing J.F. Ceratlum hyrundlnelta - ecologia unei plante complexe, mobile si de succes II Rep. Apa dulce Biol. conf. univ.
1980. N 48. P. 27-40.
75. Humphrey G.F. Pigmenți fitopiancton în Oceanul Pacific II Proc. Conf. privind măsurarea productivității primare, marine și de apă dulce, desfășurată la Univ. Hawaii, 21 august - 6 septembrie 1961. NE. Comisia pentru Energie Atomică, 1963. p. 121-141.
76. Ichimura S„ Aruga Y. Câteva caracteristici ale fotosintezei fitopianctonului de apă dulce // Bot. Mag., Tokyo. 1958. Vol. 71, N 841-842. P. 261-269.
77. Jeffrey S.W., Humphrey G.F. Noi ecuații de spectrofotometrie pentru determinarea clorofilelor „a”, b”, „ci” și „сг” în plante superioare, alge și fitopiancton natural II Biochem. Physiol. Pfianz. 1975. Bd. 167, N2. P. 191-194.
78. Jewson D.H. Interacțiunea componentelor care controlează fotosinteza net Într-un lac bine amestecat (Lough Neagh, Irlanda de Nord) II Freshwater Biol. 1976. Vol. 6, N 6. P. 551-576.
79. Jewson D.H. Pătrunderea luminii în raport cu conținutul de fitopiancton din zona eufotică a Lough Neagh, N. Ireland//Oicos. 1977. Vol. 28. N 1.P. 74-83.
80. Jewson D.H., Tailing J.F., Dring M.J., Tilzer M.M., Heaney S.I., Cunningham C. Measurement of photosynthetically available radiation in freshwater: comparative tests of some current" instruments used in studies of primary production II J. Plankton Res. 1984. 6, nr. 2. P. 259273.
81, Kirk J.T.O. Lumina si fotosinteza In ecosistemele acvatice. Cambridge. 1983. .
82. Koski V.M., Smith J.H.C. Proprietățile de izolare și absorbție spectrală ale protoclorofilei din răsaduri de orz etiolat II J. Am. Chim. 1948. Vol. 70. P. 3558-3562. ________________
83. Lorenzen C.J. Determinarea clorofilei si a feopigmentelor: ecuatii de spectrofotometrie // Llmnol. Oceanogr. 1967. Vol. 12, N 2. P. 343-346.
84. Loronzen C.J., Jeffrey S.W. Determinarea clorofilei în apa de mare II Lucrări tehnice UNESCO În științe marine. 1980, N 35 P. 3-20.
85. Manning W.M., Juday R.E. Conținutul de clorofilă și productivitatea unor lacuri din nord-estul Wisconsinului // Trans. Înţelept. Acad. Sci. 1941. Vol. 33. P. 383-392.
88. Marfwr A.F.H., Kusch E.A., Ra! H., Rfsmann 3. Tha iT-oosurcment of phctcsynthsilc pigments in freshwater and standartizatlon of methods: Concluzii si recomandari // Arh. Hydroblol., Beih.Ergebn. Llmhol. 1980. H. 14. P. 91-106.
87. Moreth C.M., Yentsch C.S. Rolul chiorofilazei și luminii în descompunerea clorofilei din fitoplanctonul marin H J. Ex>. Mar. Biol. Ecol. 1970. Vol. 4, N 3. P. 238-249.
88. Neale P.J.M. Tailing J.F., Heaney S.I., Reynolds C.S., Lund J.W.6. Serii lungi de timp din districtul englezesc al lacurilor: dinamica fitoplanctonului iradiant-spandant în timpul maximului de primăvară // Llmncl. Ccenogr. 1991. Vol. 36, N 4. P.751-760.
09. Parsons T.R., Strickland J.D.H. Discuție privind determinarea prin spectrofotometrie a pigmenților plantelor marine cu ecuații revizuite pentru determinarea clorofilelor și carotenoidelor//J. Mar. Res. 1983. Vol. 21, N3. P. 155-163.
ASA DE. Plait T., Galiegos C.L., Harrison W.G. Fotoinhlbltion de fotosinteză în ansambluri naturale de fitoplancton marin // J. Mar. Res. 1980. Vol. 38, N 4. P. 687-701.
91. Piatt T., Sathyendranath S., Ravindran P. Producția primară de fitoplancton: soluții analitice pentru rate zilnice pe unitatea de suprafață a apei//Proc. R. Soc. Londra. 1990. Vol. 241. P. 101-111.
92. Raportul acestei prime reuniuni a acestui grup comun de experți în energia radiantă fotosintetică II Lucrări tehnice UNESCO în științe marine. 1965. N 2.
93. Richards F.A., Thompson T.G. Estimarea și caracterizarea populațiilor de fitoplancton prin analize pigmentare. ft.O metodă de spectrofotometrie pentru estimarea pigmenților de plancton //J. Mar. Res. 1952. Vol. 11, N 2. P. 156-172.
94. Schomer H. Fotosinteza plantelor acvatice la diferite adâncimi în lacurile din nord-estul Wisconsinului // Ecologie. 1934. Vol. 15, N 2. P. 217218.
95. Schomer H.A., Juday C. Fotosinteza algelor la diferite adâncimi în unele lacuri din nord-estul Wisconsinului. I. Observaţii din 1933//Trans. Înţelept. Acad. Sci. 1935. Vol.29. P. 173-193.
96. Grupul de lucru SCOR-UNESCO N 17. Determinarea pigmenților fotosintetici II Determinarea pigmenților fotosintetici în apa de mare. Monografii de metodologie oceanografică. 1966.1. P. 9-18.
97. Stra^krafaa M., Gnauk A. Freshwater ecosistems - modeling and simulations. Amsterdam. 1985.
98. Szumiec M. Efectul eutroflaţiei controlate asupra radiaţiei solare care pătrunde în iazuri II Acta Hydroblol. 1975. Vol. 17, N 2. P. 149-182.(nr/ibCK.)
99. Tailing J.F. Caracteristicile fotosintetice ale unor diatomee de apă dulce În raport cu radiația subacvatică II New Phytol. 1957. Vol. 56, N. 1. P. 29-50.
100. Tailing J.F. Efecte de auto-umbrire În populațiile naturale ale unei diatomee planctonlc // Wetter u. Leben. 1960. Jahrg. 12, H.9-10. S. 235242.
101. Tailing J.F., The underwater light climate as a controlling factor in the production ecology of freshwater phytopiankton // Mitt, int. Verein, theor. angew. Llmnol. 1971, N 19. P. 214-243.
102. Tailing J.F. Utilizarea radiației solare de către fitopiancton // Trends in photobiology.N.Y. 1982. P. 619-631.
103. Tailing J.F. Schimbări sezoniere comparative și variabilitate și stabilitate interanuală, într-o înregistrare de 26 de ani a biomasei totale de fitopiancton în patru bazine ale lacului englez // Hydrobiologla. 1993. Vol. 268, N1. P. 65-98.
104. Tailing J.F., Driver D. Câteva probleme în estimarea clorofilei „a” în fitopiancton // Proc, Conf. privind măsurarea productivității primare, marine și de apă dulce, desfășurată la Univ. Hawaii, 21 august-6 septembrie 1961. Comisia pentru Energie Atomică din SUA. 1963. P. 142-146.
105. Tailing J.F., Wood R.B. Prosser M.V., Baxter R.M. Limita superioară a productivității fotosintetice de către fitopiancton: dovezi din lacurile de sodă etiopiene // Freshwater Biol. 1973. Vol. 3, N 1. P. 53-76.
106. Ttlzer M.M. Importanța absorbției fracționale a luminii de către pigmenții fotosintetici pentru productivitatea fitopianctonului în Lacul Constance II Limnol. Oceanogr. 1983. Vol. 28, N 5. P. 833-846.
107. Tilzer M.M. Randamentul cuantic ca parametru fundamental care controlează profilele fotosintetice verticale ale fitopianctonului // Arh. Hidrobiol., Suppl. 1984. Bd. 69, N 2. S. 169-198.
108. Tilzer M.M., Schwarz K. Modele sezoniere și verticale ale adaptării luminii fitoplanctonului într-un lac de munte înalt // Arh. Hidroblol. 1976. Bd. 77, N 4. 5. 488-504.
109. Thom.asson K. Reflecție asupra lacurilor arctice și alpine II Oikos. 1958. Vol. 7, N 1. P. 117-143. ____________________________
110. Vollenwieder R.A., Tailing J.F., Westlake D.F. Un manual privind metodele de măsurare a producției primare în mediile acvatice. Oxford. 1974.
111. Yentsch C.S. O metodă non-extractivă pentru estimarea cantitativă a clorofilei în culturi de alge // Nature. 1957>Vol. 179, N
4573. P. 1302-1304.
112. Yentsch C.S. Relația dintre clorofilă și producția de carbon fotosintetic cu referire la măsurarea produșilor de descompunere ai pigmenților cloroplastici H Mem. 1. Ital. Idrobiol. Supl. .13., 1255. P. 323-3“in.
Principalele publicații pe tema tezei
Pyrina I.L. Intensitatea fotosintezei la alge în legătură cu iluminarea sezonieră // Tr. Inst. biol. Rezervor 1959. T. 1(4). pp. 102-109.
2. Pyrina I.L. Produse fotosintetice în Volga și rezervoarele sale // Buletin. Ixt. biop. Rezervor 1959(a). N 3. P. 17-20.
3. Pyrnns I.L. Dependența producției primare de compoziția fitoplanctonului // Producția primară a mărilor și a apelor interioare. Minsk, 1961. P. 30.8-313.
4. Pyrina I.L. Rezultate preliminare, aplicarea metodei spectrofotometrice pentru determinarea pigmenților fitoplanctonului de apă dulce // Aspecte biologice ale studierii rezervoarelor. M.-L., 1963. P. 51-59.
5. Pyrina I.L. Fotosinteza fitoplanctonului de apă dulce în diferite condiții de lumină într-un rezervor // Rezumate ale rapoartelor. 1 întâlnire privind problemele ciclului materiei și energiei din rezervoarele lacului. Listeenichnsya din Baikal, 1964. P. 81-62.
6. Pyrina I.L. Fotointegrator subacvatic // Gidrobiol. revistă 1365, T.1, N 2. P. 61-07,
7. Pyrina I.L. Evaluarea producției primare de fitoplancton pe baza rezultatelor măsurătorilor conținutului de clorofilă și a intensității radiațiilor subacvatice // Probleme de hidrobiologie (Rezumate ale Primului Congres al Societății Botanice de Stat All-Russian). M„ 1965 (a). p. 359-360.
8. Pyrina I.L., Feoktistova O.I., Rolul algelor albastre-verzi în producerea de materie organică într-un rezervor // Ecologia și fiziologia algelor albastre-verzi. M.-L., 1965. P. 42-46.
9. Pyrina I.L. Producția primară de fitoplancton în rezervoarele Ivankovo, Rybinsk și Kuibyshev în funcție de unii factori // Producția și circulația materiei organice în rezervoarele interioare - L., 1966. P. 249-270.
10. Pyrina I.L. Producția primară a unor rezervoare în legătură cu iluminarea, clorofila și biomasa fitoplanctonică // Rezumat de doctorat. insulta. Minsk, 1966 (a). 16 pagini
11. Pyrina, I.L., Makeeva N.L. Despre metoda de determinare a clorofilei în celulele de alge colectate pe un filtru membranar // Vegetația rezervoarelor Volga. M.-L., 1966. P. 198202.
12. Pyrina IL. Fotosinteza fitoplanctonului de apă dulce în diferite condiții de lumină într-un rezervor // Ciclul materiei și energetice în rezervoarele lacului. M., 1967. S. 202-208.
13. Pyrina I.L. Eficiența utilizării energiei solare în timpul fotosintezei planctonului în rezervoarele Volga // Factori radianți în viața organismelor acvatice. L., 1967 (a). pp. 34-42.
14. Pyrina I.L. Dependența fotosintezei fitoplanctonului de biomasa și conținutul de clorofilă // Microflora, fitoplanctonul și vegetația superioară a corpurilor de apă interioare. L., 1967 (b). pp. 94-103.
15. Pyrina I.L. Producția primară în rezervoarele din Volga de Sus și Mijloc // Rezumate ale rapoartelor. eu conf. privind studiul rezervoarelor din bazinul Volga. Tolyatti, 1968. p. 88-80.
16. Pyrina I.L. Intensitatea și compoziția spectrală a luminii solare în ape optic diferite // A doua întâlnire. privind problemele ciclului materiei și energiei din rezervoarele lacului. Rezumatul rapoartelor, partea I. Listvenichnoe despre Baikal, 1969. pp. 83-84.
17. Pyrina I.L., Elizarova V.A. Conținutul de clorofilă în fitoplanctonul unor corpuri de apă dulce // A doua întâlnire. privind problemele ciclului materiei și energiei din rezervoarele lacului. Rezumatul rapoartelor, partea II. Zada pe Baikal, 1969. p. 9-10.
18. Pyrina I.L. Iluminarea subacvatică și clorofila ca indicatori ai producției primare a rezervoarelor // Procese biologice în rezervoare marine și continentale. (Tezele raportului și congresului VSBO). Chișinău, 1970. P. 314.
19. Pyrina I.L., Elizarova V.A. Determinarea spectrofotometrică a clorofilelor în culturile unor alge II Biologia și productivitatea organismelor de apă dulce. L., 1971. P. 5665.
20. Pyrina I.L., Naumova E.I. Activitatea fotosintetică a fitoplanctonului în apele reziduale ale centralei electrice din districtul de stat Konakovo // Tezi-
Sydokl. Symp. asupra influenţei apelor termocentralelor asupra hidrologiei şi biologiei corpurilor de apă. Borok, 1971. p. 52.
21. Guseva K.A., Priymachenko A.D., Pyrina I.L. Kozhova O.M., Gerasimova N.A., Kuksn M.S., Shalar V.M. Dezvoltarea algelor albastre-verzi în rezervoarele hidrocentralelor din URSS // Rezumate de rapoarte. Toate întâlnire privind utilizarea integrată a rezervoarelor. Kiev, 1971. P. 41-42.
22. Pyrina P.L., Rutksvskaya V.A., Ilyinsky A.L. Despre influența fitoplanctonului asupra pătrunderii radiației solare în apa rezervoarelor Volga II Materia organică și elementele regimului hidrologic al lacurilor de acumulare Volga. L., 1972. S. 97-106.
23. Pyrina I.L., Getsen M.V., Elizarova V.A. Unii indicatori ai productivității fitoplanctonului în tundra lacurile Kharbey // Ciclul materiei și energiei în lacuri și rezervoare (Third Soyaeshchanke.. Rezumatul rapoartelor, colecția 1). Listvenichnoa pe Baikal, 1973. p. 112-114.
24. Pyrina Sh1., Elizarova V.A., Nikolaev I.I. Conținutul de pigmenți fotosintetici din fitoplanctonul lacului Onega și semnificația acestora pentru aprecierea nivelului de productivitate al acestui rezervor II Productivitatea lacului Onega. L., 1973. S. 108-122.
25. Pyrina I.L., Naumova E.I. Intensitatea fotosintezei fitoplanctonului în apele reziduale ale centralei electrice din districtul de stat Konakovo // Buletin. Inst. biol. intern apă informam. Buletin 1973. N 17. p. 18-22.
26. Pyryaka Ya. L. Măsurători spectrale ale radiației subacvatice în intervalul de lungimi de undă fotosintetice folosind un fotointegrator cu filtre de lumină II Procese de radiație în atmosferă și pe suprafața pământului (Materiale ale a IX-a reuniune a întregii uniuni privind actinometrie). L., 197-1.P. 423- 428.
27. Pyrina I.L. Producția primară de fitoplancton în rezervoarele Volga // Rezumate ale rapoartelor. II conf. privind studiul rezervoarelor din bazinul Volga (rezumatul rapoartelor). Borok, 1974(a). pp. 20-24.
28. Pyrina I.L. Producția primară de fitoplancton (secțiunea monografiei) II Metode de studiu a biogeocenozelor corpurilor de apă interioară. M., 1975. S. 91-107.
29. Pyrina I.L. Penetrarea și compoziția spectrală a luminii solare în ape optic diferite și ciclul materiei și energiei în rezervoarele lacului. Novosibirsk, 1^75 (a). p. 349-353.
ZO^Pyrika I.L., Elizarova V.A. Conținutul de clorofilă în fitoplanctonul unor corpuri de apă dulce II Ciclul materiei și energiei în rezervoarele lacustre. Novosibirsk, 1975. P. 85-89.
31. Pyrina I.L., Elizarova V.A. Intensitatea fotosintezei și conținutul de clorofilă în forme de masă de fitoplancton în rezervoare de diferite tipuri // Studiul și protecția resurselor de apă. M., 1975 (a). p. 60.
32. Pyrina I.L., Devyatkin V.G., Elizarova V.A. Studiu experimental al efectului încălzirii asupra dezvoltării și fotosintezei fitoplanctonului // Factori antropici în viața rezervoarelor. L., 1975. S. 67-84. ,
33. Pyrina I.L., Bashkatova E.L., Sigareva L.E. Producția primară de fitoplancton în zona de mică adâncime a lacului de acumulare Rybinsk în anii 1971-1972 // Regimul hidrobiologic al apelor de mică adâncime de coastă din rezervoarele Volga Superioară. Yaroslavl, 1976. p. 106-132.
34. Pyrina I.L., Getsen M.V., Vainshtein M.B. Producția primară de fitoplancton în lacurile sistemului Kharbey al tundrei Bolshezemelskaya (secțiunea monografiei) // Productivitatea lacurilor din partea de est a tundrei Bolshezemelskaya. L., 1976. P. 63-76.
35. Elizarova V.A., Pyrina I.L., Getsen M.V. Conținutul de pigmenți de fitoplancton din apele lacurilor Kharbey.//Ibid. p. 5563.
36. Pyrina I.L., Rutkovskaya V.A. Dependența intensității fotosintezei fitoplanctonului de radiația solară totală care pătrunde în apă // Procese biologice de producție în bazinul Volga. L., 1976. P. 48-66.
37. Guseva K.A., Priymachenko-Shevchenko A.D., Kozhova O.M., Pyrina I.L., Kuksn M.S., Shalar V.M., Chaikovskaya T.S., Gerasimova N.A. Intensitatea „înfloririi” și sincronizarea vegetației în masă a algelor albastre-verzi în rezervoarele centralelor hidroelectrice ale URSS // Probleme de reglementare a „înfloririi” apei și a utilizării algelor în economia națională. Kiev, 1976. p. 6-19.
38. Pyrina I.L., Elizarova V.A., Sigareva L.E. Semne de eutrofizare a rezervorului Ivankovo în ceea ce privește productivitatea fitoplanctonului I. Eutrofizarea antropică a apelor naturale. Cernogolovka, 1977. p. 238-244.
39. Pyrina I.L. Producția primară de fitoplancton (secțiunea monografiei) // Rezervorul Ivankovo și viața sa. L., 1978. S. 102-124.
40. Pyrina I.L. Producția primară de fitoplancton (secțiunea monografiei) // Volga și viața sa. L., 1978 (a). P.148-152.
41. Pyrina I.L., Bashkatova E.L., Mineeva N.M., Sigareva L.E. Unii indicatori ai productivității fitoplanctonului în rezervorul Sheksninsky și lacurile adiacente II Resursele biologice ale corpurilor de apă din regiunea Vologda, protecția lor și utilizarea rațională
convocare (rezumate pentru o conferinţă ştiinţifico-practică). Vologda, 1978. p. 20-21.
42. Pyrina I.L., Sigareva L.E. Conținutul de pigmenți de fitoplancton din rezervorul Ivankoz în 1973-1974 // Biologia organismelor inferioare. Rybinsk, 1978, p. 3-17,
43. Pyrina I.L. Producția primară de fitoplancton în Volga (secțiunea monografie) //Râul Volga și viața sa. Haga, 1979. P. 180-194.
44. Pyrina I.L. Determinarea producției primare de fitoplancton pe baza fotosintezei maxime, radiației solare totale și transparenței apei // Gidrobiol. revistă 1979. T. 15, N 6. P. 109-113.
45. Pyrina I.L., Trifonova I.S. Cercetări privind productivitatea fitoplanctonului în Lacul Ladoga//Gidrobiol. revistă 1979. T. 15, N 4. P. 26-31.
46. Pyrina I.L., Elizarova V.A. Caracteristici comparative ale mulsurilor de diferite niveluri trofice în ceea ce privește conținutul de clorofilă // Ciclul materiei și autoepurarea biologică a corpurilor de apă. Kiev, 1980. P. 100-109.
47. Tilzer M.M., Pyrina I.L., Westlake D.F. Fotosinteza fitoplanctonului (secțiunea monografiei) // Funcționarea ecosistemelor de apă dulce. Cambridge, 1980. p. 163-170,174-176.
48. Pyrina I.L., Mineeva N.M., Korneva L.G., Letanskaya G.I. Fitoplanctonul și produsele sale (secțiunea monografiei) // Influența antropică asupra lacurilor mari din nord-vestul URSS. Partea a II-a. Hidrobiologie
și hidrochimia Lacului Alb. P., 1981. P. 15-64.
49. Pyrina I.L. Pe limita inferioară de lumină a vegetației fitoplanctonului sub gheață // Probleme de ecologie a regiunii Baikal.!!. Monitorizarea populației bacteriene a corpurilor de apă. Irkutsk, 1982. p. 85-86.
50. Pyrina I.L., Smetanin MM: Despre evaluarea valorilor medii pentru un rezervor de caracteristici cantitative ale fitoplanctonului /■ „Estimarea erorilor în studiile hidrobiologice și ihtiologice. Rybinsk, 1982, pp. 144-156.
51..Pyrina ^ I.L. . studii de radiație foto-sintetic activă care pătrunde sub gheață //Radiation Climatology and
aspecte aplicate ale actinometriei (Materials of the XI! meeting on actinometrie). Irkutsk, 1984. p. 252-254.
52, Pyrina I.L. Relația dintre radiația solară vizibilă și totală peste rezervoarele bazinului Eolga Superioară//Ciclul materiei și energiei în rezervoare. Irkutsk, 1985. p. 38-40.
53. Pyrina I.L. Condițiile regimului de lumină și dezvoltarea fitoplanctonului în perioada subglaciară în marile rezervoare lacustre din nord-vest // Probleme de cercetare a lacurilor mari ale URSS. L„ 1985 (a). pp. 111-114.
54. Pyrina I.L., Sigareva L.E. Conținutul de pigmenți de fitoplancton din rezervorul Rybinsk în ani de diferite condiții hidrometeorologice (1972-1976) // Biologia și ecologia organismelor acvatice. L., 1986. P. 65-89.
55. Mineeva N.M., Pyrina I.L. Studii ale pigmenților fitoplanctonului din rezervorul Rybinsk și Ibid. pp. 90-104.
56. Pyrina I.L., Mineeva N.M., Sigareva L.E. Studii pe termen lung ale pigmenților de fitoplancton din rezervorul Rybinsk // V Congres al VSBO, Partea I. Rezumate ale rapoartelor. Kuibyshev, 1986. p. 208-209.
57. Priymachenko A.D., Pyrina I.L., Votyakova N.E. Compoziția pigmentară și activitatea fotosintetică a planctonului și bentosului râului. Yenisei // Ibid., partea a II-a. p. 254-285.
58. Pyrina I.L., Mineeva N.M., Sigareva L.E., Balonov I.M., Mazin A.M. Despre productivitatea fitoplanctonului din Lacul Pleshcheyevo // Protecția și utilizarea rațională a apelor interne din Centrul și Nordul Câmpiei Ruse. Yaroslavl. 1986. p. 36-40.
59. Pyrina I.L. Despre slăbirea radiației solare în apele naturale II Prima Conferință Internațională Vereshchagin Baikal. Rezumate ale rapoartelor. Irkutsk, 1989. p. 35-36.
60. Pyrina I.L. Despre atenuarea radiației solare în apele naturale. „//. Prima conferință internațională Vereschagin Baikal. Rezumate. Irkutsk, 1989, p. 35-36.
61. Pyrina I.L. Caracteristici hidro-optice și regim de lumină (secțiunea monografiei) II Ecosistemul lacului Pleshcheyevo. L., 1989 (a). S. 3540."
62. Pyrina I.L., Sigareva L.E., Balonov I.M. Fitoplanctonul și capacitatea sa de producție // Ibid. pp. 71-114.
63. Pyrina I.L., Sigareva L.E., Dzyuban A.N. Producția primară de fitoplancton // Ibid. pp. 114-122.
64. Bikbulatov. E.S., Dzyuban A.N., Pyrina I.L. Bilanțul materiei organice//Ibid. pp. 216-218.
65. Dzyuban A.N., Pyrina I.L. Starea trofică și calitatea apei // Ibid. p. 218-221.
66. Pyrina I.L., Mineeva N.M. Conținutul de pigmenți de fitoplancton din coloana de apă a rezervorului Rybinsk // Flora și productivitatea fitocenozelor pelagice și litorale ale corpurilor de apă din bazinul Volga. L., 1990. p. 176-188.
67. Pyrina I.L. Dinamica pe termen lung a clorofilei și a productivității planctonului plantelor în rezervorul Rybinsk // Aspecte ecologice ale reglementării creșterii și productivității plantelor. Yaroslavl. 1991. p. 253-259.
68. Pyrina I.L. Conținutul de clorofilă și productivitatea fitoplanctonului din lac. Pleshcheyevo // Factori și procese de eutrofizare a lacului Pleshcheyevo. Yaroslavl, 1992. p. 18-21).
69. Pyrina I.L., Lyashchenko O.A. Compoziția și productivitatea fitoplanctonului în lac. Pleshcheyevo în stadiul actual. // Rezumate ale raportului. Toată rusă științific Conf., dedicată împlinirii a 300 de ani a sindicatului autohton. Pereslzl-Zalegky, 1992. p. 112-114.
70. Pyrina I.P., Lyashenko O.A. Compoziția și productivitatea fito-planetelor lacului. Pleshcheev în stadiul actual. // Tr. Toată rusă Izumi, conf. dedicat aniversării a 300 de ani a flotei ruse. yyl.Sh. Shreslavl-Zalskiy. 1392. p. 48-54.
71. Pyrina IL. Metoda oxigenului pentru determinarea producției de fitoplancton nepciv-mcfi // Aspecte metodologice în studierea producției primare de plancton în corpurile de apă interne. Syacht-Petersburg, 1053. p. 10-13.
72 Pmrina I.L. Caracteristici ale condițiilor pentru fotosinteză și probe izolate;; fitoplancton. // Ibid. pp. 21-23.
73. Pyrina PL. Determinarea actiunii fotositetice subacvatice
1IPM11L MYAP1LYAIMM N lOII WAf
74. Mbtjjwha i"i.."!., vf.-.eiaiiiiHM.fvi. Calculul valorilor integrale și medii ale producției primare și evaluarea acurateței acestora. // Ibid. pp. 30-40.
75. Pyrina IL. Analiza retrospectivă a datelor privind producția primară de fitoplancton Volga // Probleme ecologice ale bazinelor hidrografice mari. Togliatti, 1993 (a): p. 129-130.
78. Pyrina I.L., Priymachenko-A.D. Conținutul de pigmenți fito-planetoni din râu. Yenisei (secțiunea monografiei) // Studii productive și hidrobiologice ale Yenisei. Novosibirsk, 1993. P.
7-" Perina I..P., mineeza N.M., Cnrfepfetiä l.E,. .imnao;: ; A D Filimonov V.S., Mitropolskal M.U. Spatial? .."spso-fitoplanctonul copiilor (p^zdol." :> Ecologist rfi-wi, ~ FOGT.5rY prog.trlysteennogo RYSPRODSG.SIIK L.trenvshcheima (ID-roOkontov St. Petersburg, 19EZ. P. 55-8-1. 70. Pyrina I.L., Smetaniya M.M. ., Smetznnnna TL. Sgatshsh abordare a assemsvssk" thessing concentrare a pigmenților și a biomasei genei fitoplanctonului și Evaluarea productivității fitoplanctonului.Noeosibirsk, 1993. P. 30-4 4.
79. Pyrina I.L. Indicatori ai producției primare de fitoplancton în
râul rezervoare kesgzde. Volga // Microflora și penetrarea biologică în apele interioare". Sankt Petersburg (despre tipar). 60. Pyrina I.L., Dobrynin E.G., Mineeva N.M., Sigareva L.E. Despre influența fotosintezei și distrugerii materiei organice asupra regimului de oxigen al unui stratificat. lac. Ibid. 81. Pmpiche I.L., M^meeva N.M. Penetrarea luminii solare și parametrii zonei de fotosinteză în rezervorul Rybinsk // Structura și productivitatea fitoplanctonului rezervorului Rybinsk. Tolyatti (în presă)-
PESCUIT UDC 574.583(28):581 Yu. A. Gorbunova, A. V. Gorbunova * Universitatea Tehnică de Stat Astrakhan * Rezervația de Stat a Biosferei Astrakhan RELAȚIA PRODUCTIVITĂȚII FITOPLANCTONULUI DIN DELTA VOLGA CU MEDIUL FIZIC DE MEDIU FACTORI DE MEDIU FIZIC care determină factorii de mediu fizici și dinamici de sezon. fitoplanctonul, se referă în primul rând la intensitatea radiației solare și la temperatura apei. Este bine cunoscut faptul că, în procesul de fotosinteză, energia radiației solare absorbită de pigmenții fotosintetici ai fitoplanctonului, cel mai important rol dintre care îl joacă clorofila a, se transformă în energia legăturilor chimice ale compușilor organici și, prin urmare, are loc procesul de producere primară a planctonului. Alături de radiația solară, cel mai important factor care determină dezvoltarea fitoplanctonului și productivitatea acestuia este temperatura apei. Diferite tipuri de alge planctonice diferă în ceea ce privește temperatura optimă și cu cât temperatura optimă este mai ridicată, cu atât sunt capabile algele o fotosinteză mai intensă și rata de diviziune. Materiale și metode de cercetare Lucrarea se bazează pe datele de observare pe teren efectuate în zona inferioară a deltei și avandeltei Volga pe baza Rezervației Biosferei Astrakhan din 1994 până în 2004. Intensitatea fotosintezei planctonului a fost determinată printr-o modificare a oxigenului metoda balonului. Concentrația de clorofilă a a fost determinată folosind o metodă spectrofotometrică standard; calculul a fost făcut folosind formula Jeffrey și Humphrey. Sosirea radiației solare totale este prezentată conform datelor ACGMS. Pentru caracterizarea regimului de temperatură s-au folosit date de la stația de fenohidrometeo a Rezervației Biosferei Astrakhan. Prelucrarea statistică a materialului a fost efectuată folosind metode general acceptate și folosind pachete software specializate Mathcad și STATISTICA 6.0. Relația dintre productivitatea fitoplanctonului și regimul de temperatură Regimul de temperatură al cursurilor de apă și al rezervoarelor din delta Volga depinde în mare măsură de temperatura apei care intră în deltă din secțiunile din amonte ale râului și de modificările temperaturii aerului. Creșterea și scăderea temperaturii apei în cursurile de apă din zona deltei inferioare are loc treptat, fluctuația zilnică, de regulă, nu depășește un grad. Temperatura apei din stratul inferior nu diferă de temperatura stratului de suprafață - acest lucru este facilitat de un flux bun de apă și de mișcarea turbulente a curgerii. 83 ISSN 1812-9498. NEWSLETTER AGTU. 2006. Nr. 3 (32) Cursul anual al temperaturii apei determină în mare măsură diferențele sezoniere ale productivității fitoplanctonului. Factorul de temperatură devine deosebit de important în perioadele reci ale anului. Primăvara, odată cu creșterea temperaturii apei, intensitatea fotosintezei planctonului, nesemnificativă în martie, crește de 2-5 ori în aprilie. În perioada de toamnă se observă diferențe semnificative în intensitatea fotosintezei planctonului în diferiți ani, ceea ce se explică în mare măsură prin condițiile meteorologice, care pot varia foarte mult de la an la an. De regulă, atâta timp cât regiunea se află sub influența pintenului anticiclonului asiatic, vremea senină și caldă ajută la menținerea proceselor fotosintetice la un nivel ridicat. Odată cu instaurarea vremii ciclonice, însoțită de răcire bruscă și zile ploioase sau înnorate frecvente, intensitatea fotosintezei scade brusc. Regimul de temperatură al frontului deltei apei de mică adâncime diferă de regimul de temperatură al canalelor deltei de deasupra apei - stratul de apă se încălzește și se răcește mai repede. Fluctuațiile zilnice ale temperaturii apei în frontul deltei pot fi semnificative, ajungând la 10 °C. Cea mai mare denivelare în distribuția spațială a temperaturii apei se observă primăvara înainte de apariția inundațiilor, când, odată cu creșterea radiației solare și creșterea temperaturii aerului în condiții de apă mică, o încălzire semnificativă a zonei de apă din frontul deltei. apare în comparație cu cursurile de apă. În unele zone ale frontului deltei, diferența de temperatură a apei în comparație cu cursurile de apă poate fi de 10–15 C. În această perioadă, temperatura este factorul principal în eterogenitatea spațială a fitoplanctonului; intensitatea vegetației fitoplanctonului din frontul de deltă, în special în anumite zone, o depășește semnificativ pe cea din canale. Cele mai productive zone sunt zonele de coastă bine încălzite ale insulelor. Pentru cursurile de apă din zona inferioară a deltei Volga, atunci când se efectuează o analiză a corelației dintre suma temperaturilor apei timp de o lună și rata medie de fotosinteză a fitoplanctonului timp de o lună și suma temperaturilor apei timp de o lună și concentrația medie lunară de clorofila a în plancton, în ambele cazuri s-a obținut o natură diferită a relației pentru fitoplanctonul vară-toamnă și fitoplanctonul de primăvară și toamna târzie (Fig. 1). Corelația dintre suma temperaturilor apei pe lună și rata medie a fotosintezei fitoplanctonului pe lună s-a dovedit a fi pozitivă și destul de ridicată. În cazul fitoplanctonului de primăvară și toamnă târzie, coeficientul de corelație a fost de +0,90 (rmin = 0,51; р< 0,05); в случае летнеосеннего фитопланктона соответственно +0,62, (rmin = 0,42; р < 0,05). Корреляция между суммой температур воды за месяц и среднемесячной концентрацией хлорофилла а в планктоне также оказалась положительной и довольно высокой. В случае весеннего и позднеосеннего фитопланктона коэффициент корреляции составил +0,92 (rmin = 0,42; р < 0,05); в случае летне-осеннего фитопланктона соответственно +0,66 (rmin = 0,43; р < 0,05). 84 РЫБНОЕ ХОЗЯЙСТВО гО2 м-3 сут-1 мгм -3 1,8 1,6 1,4 1,2 1,0 0,8 0,6 0,4 0,2 0,0 12 10 8 6 4 2 0 0 5 1 10 215 а 20 25 30 0 10 20 оС С 30 СоС б Рис. 1. Зависимость средней за месяц интенсивности фотосинтеза (а) и среднего за месяц содержания хлорофилла а в планктоне (б) от среднемесячной температуры воды: 1 – весенний и позднеосенний фитопланктон; 2 – летне-осенний фитопланктон. Ось абсцисс – температура воды; ось ординат: а – интенсивность фотосинтеза планктона; б – содержание хлорофилла а в планктоне Для весеннего и позднеосеннего фитопланктона различие коэффициентов корреляции между температурой воды и скоростью фотосинтеза фитопланктона и между температурой воды и уровнем содержания хлорофилла а незначимо (uz`-z`` = 0,14; u05 = 1,96). В то же время для летнеосеннего фитопланктона различие между обоими выборочными коэффициентами корреляции является значимым (uz`-z`` = 2,27; u05 = 1,96). Связь продуктивности фитопланктона с солнечной радиацией Количество солнечной радиации, доходящее до поверхности воды, определяется географической широтой и изменяется с погодными условиями. В среднем за год общий приход суммарной солнечной радиации в регионе составляет 5 146 МДж·м-2. Интенсивность солнечной радиации значительно меняется по сезонам года, что связано с годовым ходом высоты Солнца. Поверхность региона получает в летние месяцы в 4,5 раза больше солнечной радиации по сравнению с зимним периодом. Погодные условия дельты Волги с середины апреля и до середины октября в основном обусловлены воздействием отрогов азорского антициклона, перемещающихся с запада на восток. При таких синоптических процессах устанавливается малооблачная погода . Солнечные лучи, падая на водную поверхность, частично отражаются от нее, частично же, преломляясь, проникают вглубь. Световой поток, проникая в воду, подвергается ослаблению за счет избирательного поглощения и рассеяния тем сильнее, чем больше содержание взвешенных частиц. Мутность воды в низовьях дельты Волги зависит как от количества взвешенных наносов, приносимых Волгой, так и от наносов, поступающих в водоемы вследствие эрозионных процессов, происходящих в руслах водотоков дельты . Максимальное содержание в воде взвесей в дельте наблюдается в первой половине мая – значительно раньше прохожде85 ISSN 1812-9498. ВЕСТНИК АГТУ. 2006. № 3 (32) ния пика половодья. В авандельте содержание в воде взвесей значительно меньше, чем в водотоках дельты – на протяжении большей части года в условиях мелководья на подавляющем большинстве станций относительная прозрачность наблюдается до дна. Различие условий проникновения солнечной радиации в воду является одним из ведущих факторов, обусловливающих большую эффективность утилизации фитопланктоном энергии солнечной радиации, падающей на водное зеркало в авандельте, где значения могут достигать 0,16-0,28 %, в водотоках эти показатели достигают лишь 0,07-0,08 % . Сезонные различия интенсивности солнечной радиации в большой мере определяют изменения продуктивности фитопланктона в течение года. Это влияние проявляется в непосредственном участии энергии солнечной радиации в фотосинтетическом процессе, составляя его энергетическую основу. В то же время солнечная радиация оказывает большое влияние на температуру воды. При этом ход температуры воды в протоках запаздывает по сравнению с интенсивностью суммарной солнечной радиации примерно на месяц (рис. 2, а). Вычисление коэффициента корреляции между месячными суммами суммарной солнечной радиации за предыдущий месяц и месячными суммами температуры воды за последующий месяц показало тесную положительную связь (рис. 2, б). Коэффициент корреляции составил +0,96 (rmin = 0,42; р < 0,05). МДжм -2 900 700 500 300 100 С Январь Февраль Март Апрель Май Июнь Июль Август Сентябрь Октябрь Ноябрь Декабрь 800 600 400 200 0 1 2 3 4 МДжм-2 900 800 700 600 500 400 300 200 100 0 0 200 400 600 800 1000 С а б Рис. 2. Годовой ход величины суммарной солнечной радиации и суммы температур воды (а) и связь этих показателей (б) за 1999 и 2000 гг.: а – первая ось ординат – годовой ход величины суммарной солнечной радиации; вторая ось ординат – сумма температур воды: 1 – месячные суммы суммарной солнечной радиации за 1999 г.; 2 – месячные суммы суммарной солнечной радиации за 2000 г.; 3 – месячные суммы температуры воды за 1999 г.; 4 – месячные суммы температуры воды за 2000 г.; б – ось абсцисс – сумма температуры воды; ось ординат – суммарная солнечная радиация 86 РЫБНОЕ ХОЗЯЙСТВО Корреляционный анализ показал, что наблюдалась тесная зависимость между месячными показателями энергии солнечной радиации и средней за месяц скоростью фотосинтеза фитопланктона в водотоках (рис. 3, а). Коэффициент корреляции составил +0,81 (rmin = 0,51; р < 0,05). Была также установлена связь между месячными показателями энергии солнечной радиации и среднемесячной концентрацией хлорофилла а в планктоне водотоков (рис. 3, б). Коэффициент корреляции составил +0,83 (rmin = 0,36; р < 0,05). гО2 м-3 сут-1 1,4 мгм-3 20 1,2 15 1,0 0,8 10 0,6 0,4 5 0,2 0,0 0 0 200 400 600 а 800 1000 0 200 400 600 800 1000 б Рис. 3. Зависимость средней за месяц скорости фотосинтеза планктона (а) и средней за месяц концентрации хлорофилла а в планктоне (б) от месячного прихода суммарной солнечной радиации. Ось абсцисс – приход суммарной солнечной радиации за месяц; ось ординат: а – скорость фотосинтеза планктона; б – концентрация хлорофилла а в планктоне Заключение Продуктивность фитопланктона и ее сезонная динамика в значительной степени определяются годовым ходом температуры воды и интенсивности солнечной радиации. Корреляционный анализ выявил значимую связь скорости фотосинтеза фитопланктона и содержания хлорофилла а с этими факторами среды. При этом получен разный характер связи продукционных характеристик летне-осеннего фитопланктона и фитопланктона весеннего и позднеосеннего с температурой воды. Для весеннего и позднеосеннего фитопланктона различие коэффициентов корреляции между скоростью фотосинтеза и температурой воды и между содержанием хлорофилла а и температурой воды незначимо, в то время как для летнеосеннего фитопланктона различие между обоими выборочными коэффициентами корреляции значимо. Температура оказывает непосредственное влияние на интенсивность физиологических процессов, в том числе на скорость фотосинтеза, и лишь косвенно определяет уровень содержания хлорофилла а. Влияние солнечной радиации на интенсивность процессов фотосинтеза имеет двоякий характер: с одной стороны, она непосредственно участвует в фотосинтезе, являясь энергетической основой этого процесса, с другой – опосредованно воздействует на уровень метаболизма фитопланктона, влияя на температуру воды. 87 ISSN 1812-9498. ВЕСТНИК АГТУ. 2006. № 3 (32) Авторы выражают искреннюю благодарность Л. М. Вознесенской за помощь и данные по солнечной радиации. СПИСОК ЛИТЕРАТУРЫ 1. Роль гидрометеорологических условий в многолетней динамике продуктивности фитопланктона во внутренних водоемах / А. С. Литвинов, И. Л. Пырина, В. Ф. Рощупко, Е. Н. Соколова // Природно-ресурсные, экологические и социально-экономические проблемы окружающей среды в крупных речных бассейнах. – М.: Медиа-Пресс, 2005. – С. 70–81. 2. Михеева Т. М. Сукцессия видов в фитопланктоне: определяющие факторы. – Минск: Изд-во БГУ, 1983. – 72 с. 3. Девяткин В. Г., Метелева Н. Ю., Митропольская И. В. Гидрофизические факторы продуктивности литорального фитопланктона: Влияние гидрофизических факторов на динамику фотосинтеза фитопланктона // Биология внутренних вод. – 2000. – № 1. – С. 45–52. 4. Сиренко Л. А. Активность Солнца и «цветение» воды // Гидробиологический журнал. – 2002. – Т. 38, № 4. – С. 3–10. 5. Alternation of factors limiting phytoplankton production in the Cape Fear River Estuary / M. A. Mallin, L. W. Cahoon, M. R. McIver et al. // Estuaries. – 1999. – Vol. 22, N 4. – P. 825–836. 6. Eppley R. W. Temperature and phytoplankton growth in the sea // Fish. Bull. – 1972. – Vol. 70. – P. 1063–1085. 7. Винберг Г. Г. Первичная продукция водоемов. – Минск, 1960. – 329 с. 8. SCOR-UNESCO Working Group N 17. Determination of photosynthetic pigments in sea water // Monographs on oceanographic methodology. – P.: UNESCO, 1966. – P. 9–18. 9. Lorenzen C. J., Jeffrey S. W. Determination of chlorophyll in sea water. – P.: UNESCO, 1980. – 20 p. (UNESCO Techn. Pap. in Mar. Sci.; 35). 10. Jeffrey S. W., Humphrey G. F. New spectrophotometric equations for determining chlorophylls a, b, c1 and c2 in higher plants algae and na tural phytoplankton // Biochem. Physiol. Pflanzen. – 1975. – Bd. 167, N 2. – S. 191–194. 11. Урбах В. Ю. Биометрические методы. – М.: Наука, 1964. – 415 с. 12. Лакин Г. Ф. Биометрия. – М.: Высш. шк., 1980. – 293 с. 13. Гланц С. Медико-биологическая статистика. – М.: Практика, 1999. – 459 с. 14. Москаленко А. В. Характеристика гидрологического режима водоемов низовьев дельты Волги // Тр. Астрахан. заповедника. – Астрахань, 1965. – Вып. 10. – С. 37–79. 15. Вознесенская Л. М., Бесчетнова Э. И. Климатические особенности и опасные явления погоды Астраханской области в ХХ веке. – Астрахань, 2002. – 111 с. 16. Москаленко А. В., Русаков Г. В. Влияние зарегулированности водного стока реки Волги на сток взвешенных наносов в рукавах ее дельты. – Деп. в ВИНИТИ № 2997 – 79 от 08.05.79. – Вестн. МГУ. Сер. географ. – 1979. – № 3. – 14 с. 17. Горбунова Ю. А. Продуктивность фитопланктона дельты Волги: Автореф. дис. … канд. биол. наук. – Борок, 2005. – 23 с. Статья поступила в редакцию 24.03.06, в окончательном варианте – 21.04.06 88 РЫБНОЕ ХОЗЯЙСТВО THE DEPENDENCE OF PHYTOPLANKTON PRODUCTIVITY OF THE DELTA OF THE RIVER VOLGA ON PHYSICAL ENVIRONMENTAL FACTORS Yu. A. Gorbunova, A. V. Gorbunova The dependence of phytoplankton productivity of channels of the delta and the avandelta of the river Volga on the water temperature and solar radiation intensity is considered in the paper. The correlation analysis has revealed a significant relation of photosynthesis rate and chlorophyll contents with these environmental factors. The authors of the article have obtained different characters of relations of production characteristics with the water temperature for summer-autumn phytoplankton and spring and late autumn phytoplankton. 89